

Внимание: 1040 следует читать как "десять в сороковой степени".
Введение
Время от времени кто-нибудь приходит к выводу, что «поскольку случайное формирование какого-либо фермента практически невозможно, следовательно, невозможен и абиогенез». Зачастую в качестве доказательства статистической невозможности жизни приводятся впечатляющие расчёты астрофизика Фреда Хойла (Fred Hoyle) или апелляция к так называемому «Закону Бореля» (Borel). Эти люди, как и Фред, совершают одну или несколько из следующих ошибок.
Ошибки вычислений креационистов, приводящие к выводу, что «абиогенез точно невозможен»
1) Они полагают вероятность формирования «современного» белка или даже целой бактерии со всеми «современными» белками результатом случайности. Хотя это — вовсе не теория абиогенеза.
2) Они предполагают, что существует установленное число необходимых для существования жизни белков с установленными для каждого белка последовательностями.
3) Они вычисляют вероятность не параллельных испытаний, а последовательных.
4) Они не понимают значения термина «вычисление вероятности».
5) Они сильно недооценивают число функциональных ферментов/рибозимов в группе произвольных последовательностей.
Я постараюсь последовательно разъяснить вам эти многочисленные ошибки и покажу, почему невозможно никаким значимым образом провести вычисление «вероятности абиогенеза».
Примордиальная протоплазматическая глобула
Согласно вычислениям, вероятность образования случайным образом белка длиною 300 аминокислот (например, такого фермента как карбоксипептидаза) составляет (1/20)300 или один шанс из 2.04×10390, что невыразимо мало. Это число увеличивается после дополнительного учета вероятностей возникновения примерно 400 аналогичных ферментов, и итоговое число получается настолько большим, что при попытке его осмыслить мозг начинает потихоньку вытекать из ушей. Тут и создается впечатление, что формирование даже самого маленького организма абсолютно невозможно. И, тем не менее, это совершенно неверно.
Во-первых, процесс формирования биологических полимеров из мономеров подчиняется законам химии и биохимии, которые определённо не случайны.
Во-вторых, неверна изначальная предпосылка: в действительности, в современных теориях абиогенеза первая «живность» полагается гораздо более простой — даже не протобактерией, или препротобактерией (которую Опарин (Oparin) назвал протобионтом [8], а Вёзе (Woese) — прогенотой [4]), а одной или несколькими простыми молекулами, вероятно, не более чем в 30-40 субъединиц длиной. Затем такие простые молекулы постепенно развились сначала в более организованные самовоспроизводящиеся системы, а затем — в простые организмы [2, 5, 10, 15, 28]. Ниже приведена иллюстрация, сравнивающая гипотетический протобионт с современной бактерией.
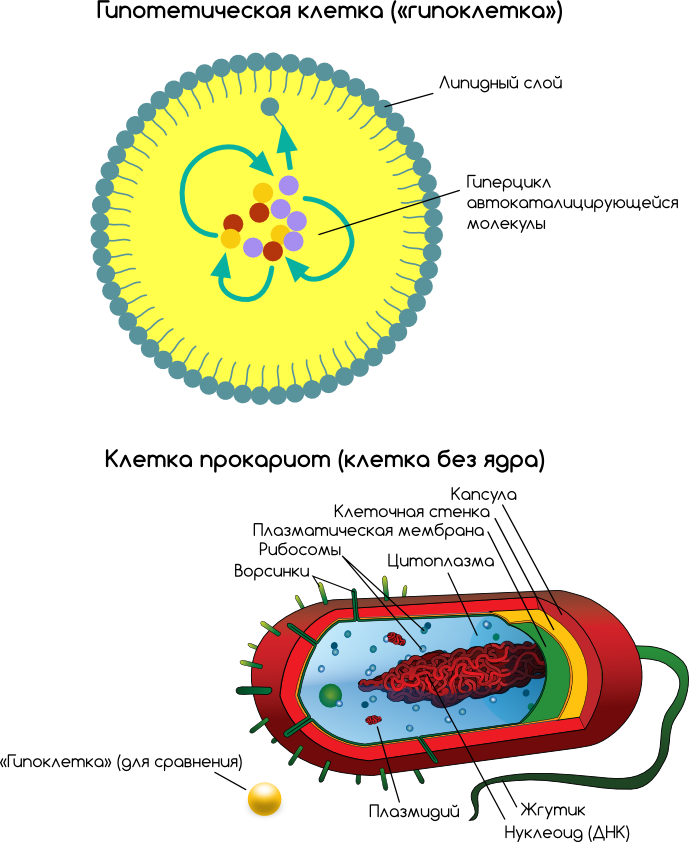
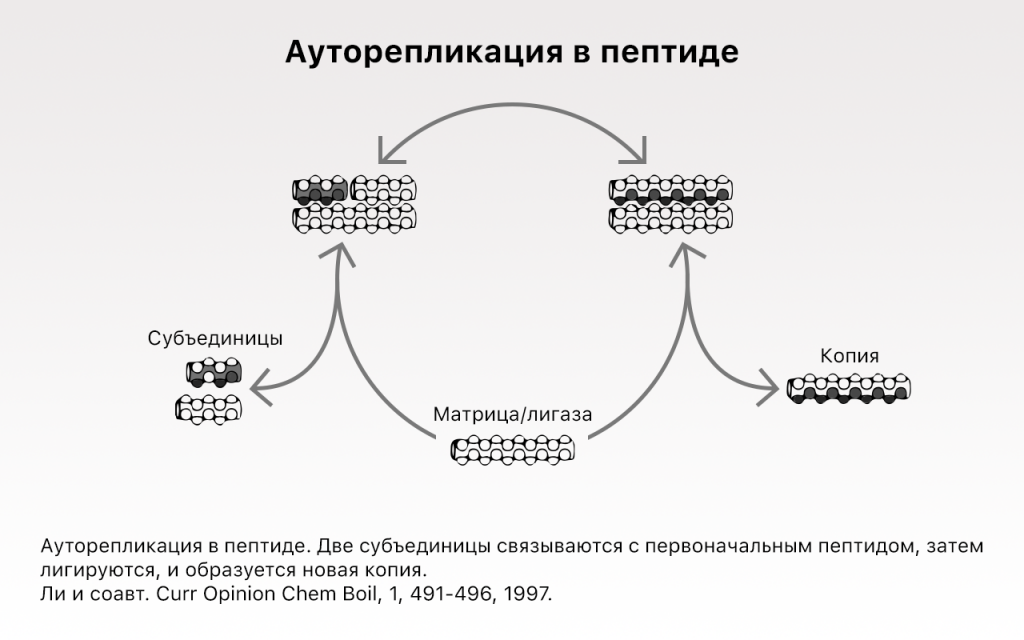
Первая «живность» могла быть одной самовоспроизводящейся молекулой, подобной «самовоспроизводящемуся» пептиду из группы Гадири [7, 17], или самовоспроизводящемуся гексануклеотиду [10], или РНК-полимеразе, воздействующей саму на себя [12].
Другая же точка зрения такова, что первыми саморепликаторами были группы катализаторов: либо белковые ферменты, либо РНК-рибозимы, регенерировавшиеся в форме каталитического цикла [3, 5, 15, 26, 28]. Примером является саморепликатор SunY из трёх субъединиц [24]. Такие каталитические циклы могут ограничиваться маленьким прудом или лагуной, либо представлять каталитический комплекс, впитанный либо глиной, либо липидными веществами на глине. Учитывая существование множества каталитических последовательностей в группе случайных пептидов или полинуклеотидов (см. ниже), не исключается, что небольшой каталитический комплекс появиться может.
Эти две модели не являются взаимоисключающими. Пептид Гадири может мутировать и образовывать каталитические циклы [9].
Независимо от того, были ли первые саморепликаторы одиночными молекулами или комплексами небольших молекул, эта модель не имеет ничего общего с хойловским (Hoyle) «торнадо, пронесшимся по свалке и собравшем Боинг-747». Чтобы ещё раз подчеркнуть свою мысль, привожу простое сравнение критикуемой креационистами теории и реальной современной теории абиогенеза.
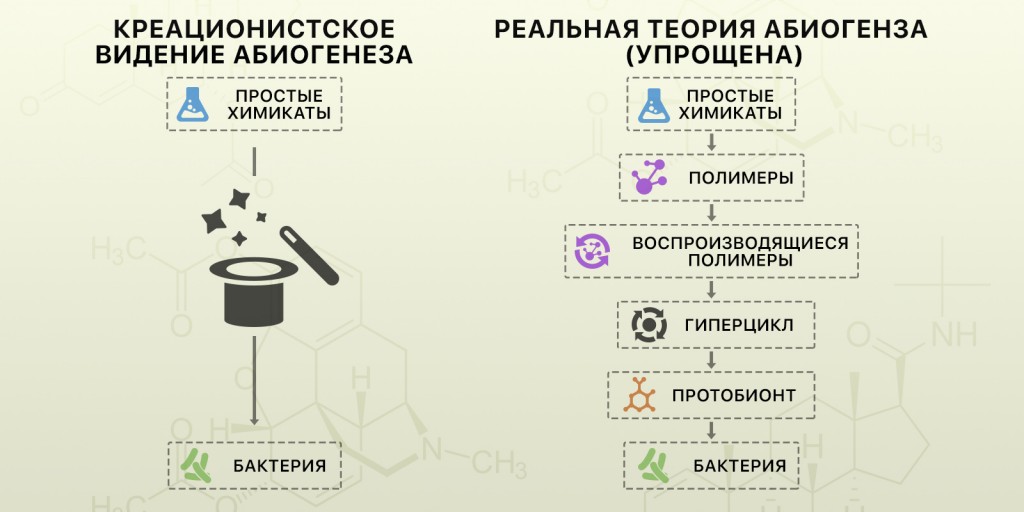
Обратите внимание, настоящая теория подразумевает ряд небольших этапов, часть которых я для простоты опустил (особенно между гиперциклом и протобионтом). Каждый этап сопровождается ростом организации и сложности. Химические компоненты усложняются до уровня организма постепенно, а не одним большим скачком [4, 10, 15, 28].
Не очень понятно, откуда креационисты взяли идею, что современные организмы сформировались спонтанно. Первая современная формулировка абиогенеза — гипотеза Опарина-Холдейна (Haldane) 20-х годов — повествует о простейших белках/протеноидах, медленно развившихся в клетки. Даже идеи 1850-х не подразумевали «спонтанности». Ближайшее же подобное прослеживается лишь в оригинальных идеях Ламарка от 1803! [8].
О чём тут можно говорить, если креационисты критикуют теорию, устаревшую более 150 лет назад и которой ни один современный эволюционный биолог не придерживается? О том, что ошибочные «опровержения» креационистов рождаются из-за наличия в статистике и биохимии некоторых фундаментальных проблем.
Миф о «жизненной цепочке»
Другое частое заявление — это что существует некая «жизненная цепочка» из 400 белков и что чтобы организмы жили, нельзя менять аминокислотные последовательности этих белков.
Однако это ерунда. Судя по всему, цифра 400 взята из генома кодирования белка бактерии Mycobacterium genetalium, обладающей наименьшим из известных в настоящее время в любом современном организме геномом [20]. Тем не менее, при ближайшем рассмотрении оказывается, что геном можно сократить до минимального генетического набора в 256 белков [20]. Ещё раз отметим, что это — современный организм. Первый протобионт/прогенота был ещё меньше [4], а предшествовали ему ещё более простые химические системы [3, 10, 11, 15].
Что касается утверждения, что последовательности белков менять нельзя, это — ещё одна бессмыслица. В большинстве белков есть регионы, где замещаема практически любая аминокислота, как и регионы, где возможны консервативные замены (когда заряженные аминокислоты можно заменить другими заряженными аминокислотами, нейтральные — другими нейтральными, а гидрофобные — другими гидрофобными). Некоторые функционально эквивалентные молекулы могут различаться аминокислотами на 30-50%. Можно даже заменить структурно неодинаковые бактериальные белки на белки дрожжей, а белки червя — на белки человека, при всём при этом организмы будут жить как ни в чём не бывало.
Так что «жизненная цепочка» — просто миф.
Подбрасывание монетки для начинающих и сборка макромолекул.
Теперь поиграем в креационистскую игру — рассмотрим формирование пептида путем случайного добавления аминокислот. Естественно, на ранней Земле пептиды возникли совершенно не так, но данное допущение будет показательным.
В качестве примера я использую «самовоспроизводящийся» пептид из вышеупомянутой группы Гадири [7]. Можно было бы взять другие примеры, такие как самореплицирующийся гексануклеотид [10], саморепликатор SunY [24] или РНК-полимераза, описанные группой Экланда (Eckland) [12], но для иллюстрации точки зрения креационистов в рамках соблюдения исторической непрерывности — небольшой пептид идеален. Длина этого пептида — 32 аминокислоты с последовательностью RMKQLEEKVYELLSKVACLEYEVARLKKVGE, являющейся также ферментом — пептидной лигазой — воспроизводящим самого себя из двух субъединиц длиною по 16 аминокислот. Кроме того, размером и составом пептидная лигаза идеально подходит для рождения путем абиотического синтеза пептидов. Тот факт, что она — саморепликатор, придает процессу дополнительной ироничности.
Вероятность её возникновения при последовательных случайных попытках равна (1/20)32, или одному шансу из 4,29х1040. Это во много-много раз вероятнее одного шанса из 2,04х10390, предлагаемого стандартным креационистским сценарием «случайного возникновения карбоксипептидазы», но всё-таки ещё очень маловероятно.
Тем не менее, есть и иной аспект оценки этих вероятностей, заключающийся в том, что большинство из нас не вполне понимает статистику. Когда нам говорят, что некое событие имеет один шанс на миллион, многие думают, что для его выпадения нужен миллион попыток, но это не так.
Следующий эксперимент вы можете проделать сами: возьмите монетку, подбросьте её четыре раза, запишите результаты и повторите всё снова. Сколько, по-вашему, раз придется повторить процедуру, прежде чем вы получите четыре орла подряд?
Вероятность выпадения четырех орлов подряд равна (1/2)4, или одному шансу из 16. Значит ли это, что прежде, чем получить четырех орлов (ОООО), мы сделаем 16 попыток? Нет: в последовательных экспериментах я проделал 11, 10, 6, 16, 1, 5 и 3 попытки прежде, чем получил ОООО. Цифра 1/16 (или 1 на миллион, или 1 на 1040) описывает вероятность события в отдельно взятой попытке, но не описывает того, как процесс будет развиваться в серии. Можно выбросить ОООО с первой же попытки (у меня получалось). Даже при одном шансе из 4,29*1040 саморепликатор может зародиться очень быстро. Но это ещё не всё.
Один шанс из 4,29*1040 — это всё равно ужасно, просто чрезвычайно маловероятно; это число трудно даже представить. Даже с учётом вышеприведенного аргумента (о том, что можно получить желаемое с первой попытки), многие скажут: «так или иначе, чтобы случайными методами получить этот репликатор, потребуется больше времени, чем существует Земля». На самом деле, нет; ведь в приведённых выше примерах мы рассматривали последовательные попытки как если бы за каждую попытку собирался только один белок/ДНК/проторепликатор. А в действительности, в процессе взаимодействия миллиардов молекул элементарных компонентов в океанах или на тысячекилометровых береговых линиях, могущих предоставить каталитические поверхности или матрицы [2,15], одновременно осуществлялись миллиарды попыток.
Давайте вернёмся к нашему примеру с монетками. Допустим, подбрасывание монетки 4 раза займёт не больше минуты; чтобы выпало ОООО, в среднем потребуется 8 минут. Теперь позовите 16 друзей, каждому дайте монету, и пусть одновременно подбросят монетки 4 раза подряд; теперь среднее время получения ОООО составит 1 минуту; теперь попробуйте выбросить 6 орлов подряд; Вероятность этого — (1/2)6, или 1 из 64. Это заняло бы в среднем полчаса, но найдите в помощь 64 человек — и получите нужный результат за минуту. Если хотите получить последовательность, имеющую вероятность 1 к миллиарду, попросите население Китая подкидывать для вас монетки и получите комбинацию в кратчайшие сроки.
Таким образом, если на нашей пребиотической Земле одновременно появлялись миллиарды пептидов, то предполагаемое время, необходимое для создания нашего репликатора, существенно сокращается.
Хорошо, теперь снова посмотрите на число 1 из 4,29×1040, это большое число, и хотя миллиард исходных молекул — много, можно ли в принципе получить достаточно молекул, чтобы случайно собрать первый репликатор менее чем за полмиллиарда лет?
Да, один килограмм аминокислоты аргинина содержит 2,85×1024 молекул (это гораздо больше миллиарда миллиардов); тонна аргинина содержит 2,85×1027 молекул. Если вы возьмете нагруженный всеми нужными аминокислотами грузовик и сбросите его в озеро среднего размера, этих молекул будет достаточно, чтобы в течение нескольких десятков лет получить наш репликатор, учитывая, что белки длиною 55 аминокислот создаются за одну-две недели [14,16].
Какое отношение всё это имеет к пребиотической Земле? Скорее всего, на ранней Земле океан имел объём 1×1024 литров. Имея аминокислоту с концентрацией 1×10-6 М (умеренно разбавленный первичный бульон, см. Chyba и Sagan 1992 [23]), имеем примерно 1×1050 потенциальных стартовых цепочек. Приличное количество работающих пептидных лигаз (около 1х1031) можно получить менее чем за год, не говоря уже о миллионах лет. Даже при вероятности 1 из 4,29х1040 синтез примитивного саморепликатора может случиться относительно быстро (помните, что наш репликатор может синтезироваться и при самой первой попытке).
Предположим, что формирование последовательности занимает неделю [14,16]. Тогда лигазу Гадири можно получить за неделю, а любая последовательность цитохрома С может возникнуть немного более чем за миллион лет (как и примерно половина всех возможных 101 пептидных последовательностей, большая часть которых будет функциональными в той или иной степени белками).
Хотя я использовал в качестве примера лигазу Гадири, те же расчёты применимы и к саморепликатору SunY или РНК-полимеразе Экланда. Оставлю это в качестве упражнения для читателя, но общий вывод (что можно получать много вещей в течение короткого времени) для этих олигонуклеотидов един.
Области поиска, или сколько иголок в стоге сена?
Итак, я продемонстрировал, что получение небольшого отдельно взятого фермента — не так уж невообразимо сложно, как креационисты (и Фред Хойл) пытаются нас убедить. Есть и ещё одно недопонимание: большинство людей полагает, что такое множество ферментов/рибозимов, не говоря уже о рибозимных РНК-полимеразах или любой форме саморепликаторов, представляет собой очень маловероятную конструкцию и что шансы формирования в результате случайной компоновки аминокислот/нуклеотидов даже одного фермента/рибозима, не говоря уже об их серии, очень малы.
Тем не менее, анализ Экланда предполагает, что во множестве всех 220-нуклеотидных последовательностей РНК целых 2,5×10112 последовательностей представляют собой работающие лигазы [12]. Что неплохо для единицы, раньше считавшейся просто структурной. Возвращаясь к нашему примитивному океану объёмом 1х1024 литров и полагая концентрацию нуклеотидов равной 1х10-7 М [23], имеем примерно 1х1049 потенциальных нуклеотидных цепей, так что приличное количество работающих РНК-лигаз (около 1х1034) могло получиться в течение года, не говоря уже о миллионе лет. Потенциальное количество РНК-полимераз тоже высоко — приблизительно одна из каждых 1020 последовательностей будет РНК-полимеразой [12]. Аналогичные соображения применимы и к синтезу рибосомных ацилтрансфераз (около одной на каждые 1015 последовательностей) и к синтезу рибозимных нуклеотидов [1, 6, 13].
Точно так же из 1х10130 возможных 100-компонентных белков только 3,8х1061 представляют цитохром С! [29] В пептидной/нуклеотидной области поиска функциональные ферменты присутствуют во множестве, так что в первичном бульоне ранней Земли весьма могла вариться работающая группа ферментов.
Таким образом, даже при более реалистичных (хотя и умопомрачительных) цифрах случайная сборка аминокислот в «жизненные» системы (и для гиперциклов на основе белковых ферментов [10], и для систем РНК-мира [18], и для сосуществующих рибозимно-белковых ферментов РНК [11, 25]) кажется вполне возможной, даже при пессимистичных показателях концентраций исходного мономера [23] и периодов синтеза.
Выводы
Неверна сама предпосылка вероятностных расчётов креационистов, так как она происходит из неправильной теории. Более того, она часто утопает в статистических и биологических ошибках.
Так как на данный момент у нас нет ни малейшего представления о том, какова вероятность зарождения жизни, практически невозможно вычислить каких-либо осмысленных вероятностей для какого-либо этапа возникновения жизни кроме первых двух (для этапа перехода от мономеров к полимерам: р=1,0, для этапа формирования каталитических полимеров: р=1,0). Для этапа от реплицирующихся полимеров к гиперциклу вероятность вполне может быть 1,0 в случае, если Кауфман прав насчёт каталитического замыкания и своих моделей фазового перехода, но для подтверждения этого требуется серьёзное углубление в химию и более детальное моделирование. Для этапа перехода от гиперцикла к протобионту вероятность зависит от теоретических концепций, находящихся в стадии разработки, и поэтому неизвестна.
Но, в конце концов, возможность возникновения жизни зависит не от бросания монетки, а от химии и биохимии, которые мы пока изучаем.
Литература
[1] Unrau PJ, and Bartel DP, RNA-catalysed nucleotide synthesis. Nature, 395: 260-3, 1998.
[2] Orgel LE, Polymerization on the rocks: theoretical introduction. Orig Life Evol Biosph, 28: 227-34, 1998.
[3] Otsuka J and Nozawa Y. Self-reproducing system can behave as Maxwell’s demon: theoretical illustration under prebiotic conditions. J Theor Biol, 194, 205-221, 1998.
[4] Woese C, The universal ancestor. Proc Natl Acad Sci USA, 95: 6854-6859.
[5] Varetto L, Studying artificial life with a molecular automaton. J Theor Biol, 193: 257-85, 1998.
[6] Wiegand TW, Janssen RC, and Eaton BE, Selection of RNA amide synthases. Chem Biol, 4: 675-83, 1997.
[7] Severin K, Lee DH, Kennan AJ, and Ghadiri MR, A synthetic peptide ligase. Nature, 389: 706-9, 1997.
[8] Ruse M, The origin of life, philosophical perspectives. J Theor Biol, 187: 473-482, 1997.
[9] Lee DH, Severin K, Yokobayashi Y, and Ghadiri MR, Emergence of symbiosis in peptide self-replication through a hypercyclic network. Nature, 390: 591-4, 1997.
[10] Lee DH, Severin K, and Ghadri MR. Autocatalytic networks: the transition from molecular self-replication to molecular ecosystems. Curr Opinion Chem Biol, 1, 491-496, 1997.
[11] Di Giulio M, On the RNA world: evidence in favor of an early ribonucleopeptide world. J Mol Evol, 45: 571-8, 1997.
[12] Ekland EH, and Bartel DP, RNA-catalysed RNA polymerization using nucleoside triphosphates. Nature, 383: 192, 1996.
[13] Lohse PA, and Szostak JW, Ribozyme-catalysed amino-acid transfer reactions. Nature, 381: 442-4, 1996.
[14] Ferris JP, Hill AR Jr, Liu R, and Orgel LE, Synthesis of long prebiotic oligomers on mineral surfaces [see comments]. Nature, 381: 59-61, 1996.
[15] Lazcano A, and Miller SL, The origin and early evolution of life: prebiotic chemistry, the pre- RNA world, and time. Cell, 85: 793-8, 1996.
[16] Ertem G, and Ferris JP, Synthesis of RNA oligomers on heterogeneous templates. Nature, 379: 238-40, 1996.
[17] Lee DH, Granja JR, Martinez JA, Severin K, and Ghadri MR, A self-replicating peptide. Nature, 382: 525-8, 1996.
[18] Joyce GF, Building the RNA world. Ribozymes. Curr Biol, 6: 965-7, 1996.
[19] Ishizaka M, Ohshima Y, and Tani T, Isolation of active ribozymes from an RNA pool of random sequences using an anchored substrate RNA. Biochem Biophys Res Commun, 214: 403-9, 1995.
[20] Mushegian AR and Koonin, EV, A minimal gene set for cellular life derived by comparison of complete bacterial genomes. Proc. Natl. Acad. Sci. USA, 93: 10268-10273.
[21] Ekland EH, Szostak JW, and Bartel DP, Structurally complex and highly active RNA ligases derived from random RNA sequences. Science, 269: 364-70, 1995.
[22] Breaker RR, and Joyce GF, Emergence of a replicating species from an in vitro RNA evolution reaction. Proc Natl Acad Sci U S A, 91: 6093-7, 1994.
[23] Chyba C and Sagan C, Endogenous production, exogenous delivery and impact-shock synthesis of organic molecules: an inventory for the origins of life. Nature, 355: 125-32., 1992.
[24] Doudna JA, Couture S, and Szostak JW, A multisubunit ribozyme that is a catalyst of and template for complementary strand RNA synthesis. Science, 251: 1605-8, 1991.
[25] Lahav N, Prebiotic co-evolution of self-replication and translation or RNA world? J Theor Biol, 151: 531-9, 1991.
[26] Stadler PF, Dynamics of autocatalytic reaction networks. IV: Inhomogeneous replicator networks. Biosystems, 26: 1-19, 1991.
[27] Eigen M, Gardiner W, Schuster P, and Winkler-Oswatitsch R, The origin of genetic information. Sci Am, 244: 88-92, 96, et passim, 1981.
[28] Eigen M, and Schuster P, The hypercycle. A principle of natural self-organization. Springer-Verlag, isbn 3-540-09293, 1979.
[29] Yockey HP, On the information content of cytochrome c. J Theor Biol, 67: 345-76, 1977.
Авторы: Ян Масгрэйв, Lies, Damned Lies. Источник: Statistics and Probability of Abiogenesis Calculations.
Над переводом работали: Анастасия Ткаченко, Олег Коптев, Дарья Чернышева, Александра Резайкина, оформили Полина Бальцевич и Радмир Абильев.
Дополнительно:
Марков Александр. Проблемы происхождения жизни. Абиогенез.
Еськов Кирилл. Происхождение жизни на Земле.
Никитин Михаил. Зарождение жизни на Земле и других планетах.
Возникновение жизни (рассказывает биолог Михаил Никитин)
Происхождение жизни на Земле (рассказывает биолог Михаил Никитин)
Начало эволюции на Земле. Как неживое вещество превратилось в живое.
Александр Марков. Происхождение жизни, начало эволюции на Земле.
Расширение белковой вселенной продолжается.
Сколько у вас хромосом? История одной мутации.
Бактериальные жгутики неуменьшаемо сложны.
Новое в теории появления жизни.








Оценили 15 человек
26 кармы