
В целях ликбеза в рядах "стихийных" антиэволюционистов (т.е. не идейных - неизлечимых, а просто недоучившихся, с которыми не всё ещё потеряно) публикую на Конте отличную книгу "Доказательства эволюции" (2010 г., авторы: Борисов Н. М., Воробьев Ф. Ю., Гиляров А. М., Еськов К. Ю., Журавлев А. Ю., Марков А. В., Оскольский А. А., Петров П. Н., Шипунов А. Б. Под редакцией А.В.Маркова.). Поскольку её объём довольно велик, и мало кто осилит его сразу и целиком, публикую книгу по частям. Каждая из них представляет интерес. Сама книга (в HTML) доступна по ссылке. Желающие читать её в форматах FB2, ePub и др. могут найти соответствующие ссылки при помощи поисковых сервисов.
Группы доказательств:
6.Молекулярно-генетические и биохимические доказательств
Открытия молекулярной генетики блестяще подтвердили факт эволюции
Дарвин опубликовал «Происхождение видов» почти за 100 лет до расшифровки структуры ДНК. Новые знания, полученные с тех пор, могли бы однозначно опровергнуть эволюционное учение, если бы оно было ложно. Вместо этого анализ ДНК дает нам убедительнейшие доказательства теории эволюции. Сам факт наличия наследственной изменчивости необходим для эволюции, и если бы оказалось, что ДНК устойчива к изменениям, это означало бы конец теории. Но ДНК постоянно мутирует, порождая генетическое разнообразие, которое служит материалом для отбора. При этом обычно чем больше генетических различий между организмами, тем сильнее различается и их строение (хотя это не строгое правило, поскольку многие генетические мутации не проявляются в фенотипе). Например, отличия генома человека от генома шимпанзе включают 35 миллионов замен отдельных нуклеотидов, 5 миллионов удалений и вставок, слияние двух хромосом и девять хромосомных инверсий. Это очень небольшая степень различия (порядка 1-2%), учитывая, что размер генома человека и шимпанзе - свыше 3 миллиардов пар нуклеотидов. Все типы мутаций, которые привели к возникновению этих различий, наблюдаются и сегодня у разных организмов как в природе, так и в лаборатории; в противном случае версию об эволюционном происхождении от общего предка пришлось бы пересматривать, то есть это еще один пример фальсифицируемости теории эволюции.
Расшифровка молекулярной основы наследственности (ДНК) и генетического кода на самом деле была важнейшим "моментом истины" в истории эволюционного учения. "Вещество наследственности" вполне могло оказаться разным у разных видов (например, у человека - ДНК, а у шимпанзе - какой-нибудь другой биополимер). Генетический код тоже мог оказаться разным. В обоих случаях эволюционное превращение одного вида в другой, как и их происхождение от общего предка, стало бы принципиально невозможным, и эволюционная теория была бы опровергнута. Но и "вещество наследственности" (полинуклеотиды ДНК и РНК), и генетический код оказались одинаковыми у всех без исключения форм жизни - от вирусов и бактерий до человека включительно. Правда, в генетическом коде изредка встречаются вариации (см.: Генетический код допускает разночтения), но они очень невелики и обычно затрагивают только некоторые второстепенные, "избыточные" кодоны. Эволюционная теория четко объясняет, почему генетический код практически не может изменяться в ходе эволюции. Чисто "технически" радикальное изменение генетического кода осуществить легко и просто: достаточно внести несколько десятков мутаций в гены транспортных РНК - молекул, играющих ключевую роль в "считывании" кода. В результате, например, "триптофановая" тРНК, распознающая кодон УГГ и присоединяющая к синтезируемой молекуле белка аминокислоту триптофан, начнет распознавать другой кодон или кодоны, например АГГ и АГА, которые сейчас кодируют аргинин. Но в результате этой простой мутации произойдет радикальное изменение всех белков, синтезируемых клетками организма: во всех белковых молекулах там, где должен быть аргинин, окажется триптофан. Такое изменение, затронувшее сразу все белки, не может не оказаться чрезвычайно вредным для организма. Соответственно, такая мутация немедленно будет отсеяна отбором. Антиэволюцинизм, напротив, не может предложить никаких внятных объяснений наблюдаемого единства генетического кода у всех организмов. Творец вполне мог бы снабдить разные виды сотворенных им существ разными генетическими кодами - ну хотя бы для того, чтобы не вводить биологов во искушение, предоставляя им еще один чрезвычайно весомый довод в пользу эволюции. К тому же это было бы и полезно для организмов, так как предотвратило бы межвидовую передачу болезнетворных вирусов. Именно таким путем человек "обзавелся" вирусами оспы (от рогатого скота), СПИДа (от обезьян), гриппа и др. Чисто "технически" разные варианты кода совершенно равнозначны и работать смогли бы одинаково хорошо.
Различия между геномами видов должны соответствовать не только экспериментально наблюдаемым типам мутаций, но и филогенетическому дереву, и палеонтологической летописи. Подобно тому, как анализ ДНК позволяет установить степень родства между двумя людьми, тот же самый анализ ДНК (сравнение отдельных генов или целых геномов) позволяет выяснить степень родства между видами, а зная количество накопленных различий, исследователи определяют время расхождения двух видов, то есть время, когда жил их последний общий предок. Например, согласно данным палеонтологии, общий предок человека и шимпанзе жил примерно 6 миллионов лет назад (такой возраст имеют, например, ископаемые находки оррорина и сахелантропа - форм, морфологически близких к общему предку человека и шимпанзе). Для того, чтобы получилось наблюдаемое число различий между геномами, на каждый миллиард нуклеотидов должно было приходиться в среднем 20 изменений за одно поколение. Сегодня у людей скорость мутаций составляет 10-50 изменений на каждый миллиард нуклеотидов за одно поколение (Giannelli, F., Anagnostopoulos, T., Green, P. M. Mutation rates in humans. II. Sporadic mutation-specific rates and rate of detrimental human mutations inferred from hemophilia B.), то есть данные палеонтологии согласуются с результатами анализа ДНК, в строгом соответствии с теорией эволюции (см. материал Genetic rates of change из архива TalkOrigins.org.)
Для того, чтобы построить филогенетическое дерево, достаточно рассмотреть несколько генов, присутствующих у всех организмов, которые мы хотим включить в это дерево (обычно чем больше генов, тем статистически достовернее получаются элементы дерева - порядок ветвления и длины ветвей).
Особый интерес представляют случаи, когда различия геномов оказываются нейтральными, то есть не влияют на организм. Например, было рассчитано, что цитохром c может быть составлен как минимум 2.3 * 10^93 разными способами за счет того, что одинаковую по функции и биологически значимым свойствам молекулу белка можно получить с помощью разных последовательностей аминокислот. В свою очередь, каждая из этих последовательностей может быть закодирована 10^46 различными последовательностями ДНК вследствие избыточности генетического кода (разные тройки нуклеотидов кодируют одну и ту же аминокислоту). Нет никаких априорных причин, кроме происхождения от общего предка, по которым два разных вида должны были бы иметь хотя бы отдаленно похожие последовательности ДНК для кодирования нормально работающего (функционального) цитохрома c. То же самое справедливо и для других белков. Тем не менее аминокислотные последовательности большинства белков у близкородственных видов (например, у шимпанзе и человека), как правило, очень похожи. Так, подавляющее большинство гомологичных белков человека и шимпанзе различаются лишь на 1-2 аминокислоты или не различаются вовсе. Различий в нуклеотидных последовательностях обычно больше за счет незначимых, или синонимичных (не влияющих на аминокислотную последовательность белка) нуклеотидных замен.
По соотношению несинонимичных и синонимичных нулеотидных замен (dN/dS) можно определить, насколько сильно действует на данный ген "очищающий" отбор, отбраковывающий мутации, которые меняют свойства белка. Как правило, чем консервативнее (постояннее) функция белка, тем ниже этот показатель. Повышение dN/dS свидетельствует о положительном отборе, т.е. о закреплении полезных мутаций. Например, повышенное значение dN/dS у человека по сравнению с другими млекопитающими зафиксировано в гене FOXP2, который связан со способностью к произнесению членораздельных звуков (см.: Будут ли расшифрованы генетические основы разума?; «Ген речи» FOXP2 оказался регулятором высокого уровня).
Малое число различий в аминокислотных последовательностях белков у близких видов связано не только с тем, что эти различия еще не успели накопиться, но и с тем, что многие одинаково удачные для выполнения данной функции аминокислотные последовательности (см. выше) отделены друг от друга так называемыми "ямами в ландшафте приспособленности". Это значит, что для того, чтобы перейти от одной такой последовательности к другой, функционально равнозначной, нужно приобрести сразу несколько мутаций, каждая из которых по отдельности может снижать функциональность белка. Многие из этих "ям" можно обойти, последовательно приобретая ряд нейтральных мутаций, но это долгий процесс, основанный на случайностях, а не на позитивном отборе, и поэтому он занимает много времени (см.: The Molecular Sequence Evidence из архива TalkOrigins.org.)
Пример сравнения нуклеотидных и аминокислотных последовательностей человека и шимпанзе
Сравнительный анализ нуклеотидных последовательностей позволяет судить о степени родства сравниваемых организмов. Это обстоятельство широко применяется на практике (в частности, для установления отцовства). Например, недавно на основе анализа ДНК из человеческих костей, обнаруженных под Екатеринбургом, удалось доказать, что это останки семьи последнего российского императора Николая II. При этом для сравнения был использован генетический материал ныне живущих родственников царской семьи (см.: Генетический анализ показал, что из детей Николая II не спасся никто).
Изучая семьи с известной генеалогией, генетики оценивают скорость накопления различий в ДНК. В частности, большую помощь оказало исследование ДНК населения Исландии - уникальной страны, где каждый житель знает всех своих предков вплоть то первых колонистов, прибывших в Исландию из Норвегии в IX веке (причем из останков нескольких первопоселенцев тоже удалось извлечь ДНК для анализа). Теми же методами можно реконструировать историю целых народов или, к примеру, находить среди современных азиатов потомков Чингисхана. Результаты генетического анализа при этом хорошо согласуются с сохранившимися историческими сведениями. В ходе многочисленных исследований такого рода, где можно было непосредственно сравнить генетические данные с историческими, генетики раз за разом убеждались в достоверности оценок родства на основе сравнения ДНК, а используемые методы развивались и совершенствовались.
Поэтому сегодня мы имеем возможность при помощи этих многократно проверенных и "откалиброванных" методов оценивать степень родства и таких организмов, по которым у нас нет прямых исторических данных. Результаты таких исследований позволяют устанавливать степень родства различных видов живых организмов с такой же степенью надежности, как и в случае установления отцовства или идентификации останков царской семьи. В частности, наше близкое родство с шимпанзе записано в наших геномах, можно сказать, аршинными буквами.
Рассмотрим пример сравнения нуклеотидных последовательностей ДНК и аминокислотных последовательностей белка у человека и шимпанзе.
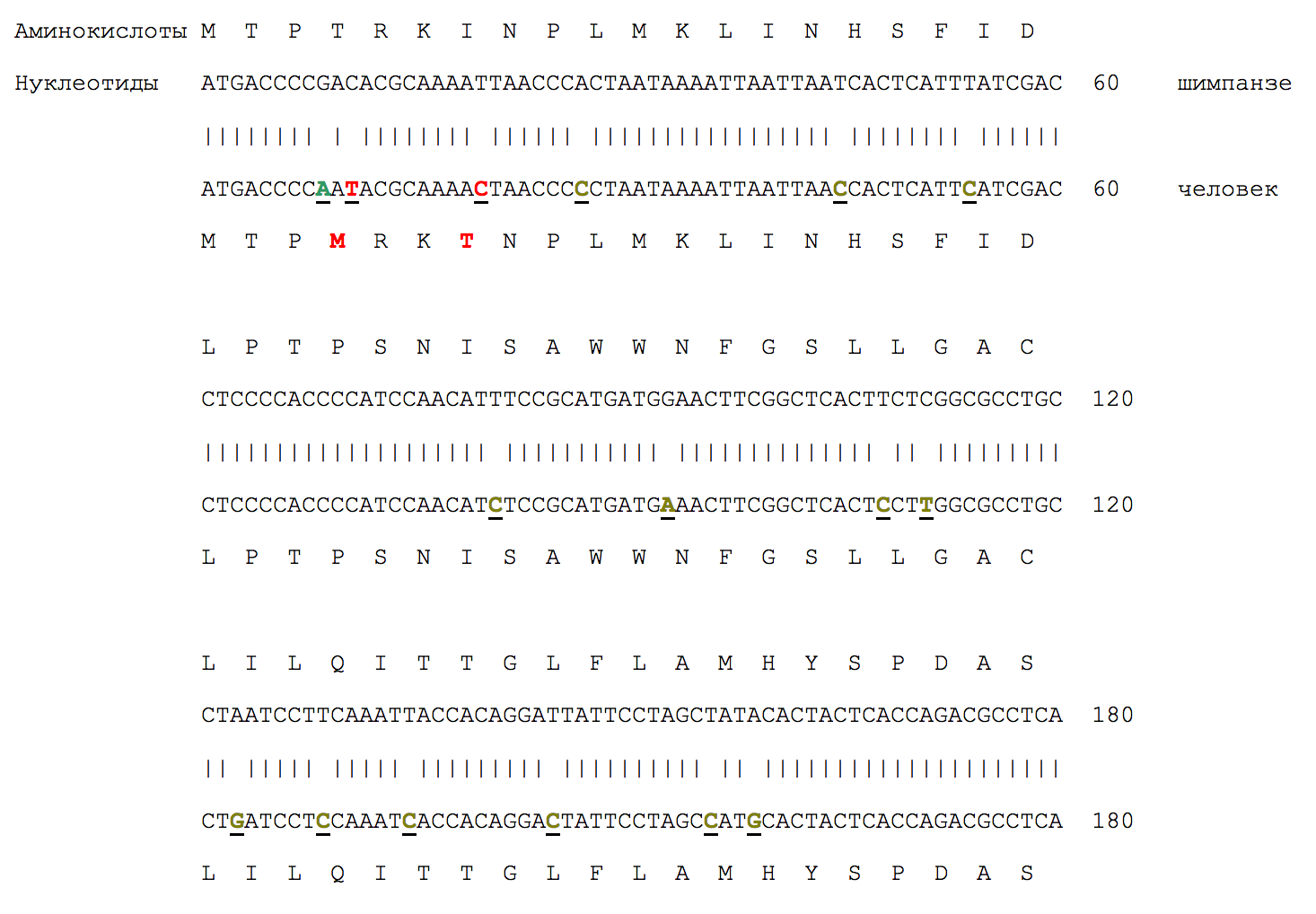
Здесь показан начальный фрагмент (180 нуклеотидов) митохондриального гена цитохрома b шимпанзе и человека. Митохондриальные гены накапливают мутации примерно в 5-10 раз быстрее, чем ядерные. Поэтому митохондриальные гены человека и шимпанзе различаются на 9%, а ядерные - где-то на 1-2%. Здесь показан митохондриальный ген, потому что если бы мы взяли ядерный ген, сходство было бы очень большим, и нам пришлось бы приводить гораздо более длинную последовательность, чтобы наглядно продемонстрировать характер различий. А вообще ген можно взять практически любой - картина будет качественно одна и та же.
Из 60 аминокислот, кодируемых этими 180 нуклеотидами, у шимпанзе и человека различаются только две (4-я и 7-я, выделены красным). Из 60 кодонов (триплетов, троек нуклеотидов) различаются 16, однако только 2 из 16 различий являются "значимыми" (несинонимичными), а остальные - синонимичные, не влияющие на структуру белка. Синонимичные нуклеотидные отличия человека от шимпанзе веделены зеленым цветом и подчеркиванием, несинонимичные - красным цветом и подчеркиванием.
Генетическое родство человека и шимпанзе доказывается даже не столько сходством последовательностей, сколько характером различий между ними. Легко заметить, что характер этих различий полностью соответствует предсказаниям эволюционной теории. Больше всего должно быть синонимичных нуклеотидных замен, потому что такие замены не влияют на свойства белка и, следовательно, невидимы для отбора, не отбраковываются им. Именно это мы и наблюдаем.
То, что в большинстве случаев (44 из 58) для кодирования одной и той же аминокислоты в геноме человека и шимпанзе используется один и тот же триплет - это еще одно доказательство генетического родства. С точки зрения функциональности нет абсолютно никакой разницы, каким из нескольких триплетов, соответствующих данной аминокислоте, закодировать ее в каждом конкретном случае. Например, аминокислота T (треонин) кодируется любым из четырех кодонов: ACA, ACT, ACG, ACC. Эта аминокислота встречается в одинаковых позициях в рассматриваемом фрагменте белка человека и шимпанзе четырежды. При этом в каждом из четырех случаев она закодирована у обоих видов одним и тем же кодоном (в первых трех случаях это кодон ACC, в четвертом - ACA). Вероятность случайности такого совпадения 0.25^4 = 0.0039. Если собрать все такие случаи по геномам человека и шимпанзе, вероятность случайности получится невообразимо ничтожной, практически неотличимой от нуля.
Таким образом, дело здесь не просто в сходстве ДНК, дело в характере сходства, которое выходит далеко за пределы любой функциональной оправданности. Особенно важно сходство по бессмысленным частям генетического "текста" (сюда относится и использование одинаковых синонимичных кодонов), а также по характерным ошибкам в нем (см. ниже об эндогенных ретровирусах и псевдогенах). Для любого специалиста по сравнительной геномике кровное родство человека и шимпанзе абсолютно очевидно и не вызывает даже тени сомнения. Опытный учитель сразу поймет, что один ученик бездумно списал у другого, если заметит в их сочинениях не только одинаковые мысли (это еще можно объяснить одинаковыми намерениями авторов), но и одинаковые фразы, используемые для их выражения, а особенно - одинаковые ошибки и одинаковые сорные словечки в одних и тех же местах текста. Все эти бесспорные признаки единства происхождения (а не независимого сотворения) в величайшем изобилии присутствуют в геномах близкородственных видов, каковыми являются человек и шимпанзе.
Сравним теперь аминокислотные последовательности того же самого фрагмента цитохрома b у шимпанзе, человека и макаки резуса:

Как видим, у макаки аминокислотная последовательность этого белка сильнее отличается от человеческой и шимпанзиной, чем последовательности первых двух видов друг от друга (14 аминокислотных различий между макакой и шимпанзе, 13 - между макакой и человеком, 2 - между шимпанзе и человеком). Это полностью соответствует биологической систематике и эволюционному дереву (шимпанзе - гораздо более близкий родственник человека, чем макака). То, что по одной аминокислоте (4-й) макака больше похожа на человека, чем на шимпанзе, означает, что, скорее всего, у общего предка макаки и человекообразных в этой позиции стояла аминокислота M, которая сохранилась у макаки и человека. Однако в линии шимпанзе, уже после ее отделения от человеческой линии, произошла замена M на T.
Интересно взглянуть на ситуацию с точки зрения шимпанзе. Для этого вида человек - более близкий родственник, чем любая другая обезьна. Даже горилла, внешне не так уж сильно отличающаяся от шимпанзе (по крайней мере на наш человеческий взгляд), приходится шимпанзе более дальней родственницей, чем человек. В свою очередь, для гориллы люди и шимпанзе - самые близкие родственники, значительно более близкие, чем любые другие обезьяны.
Таким образом, результаты сравнения генов и белков подтверждают представления о родственных связях между видами (эволюционном дереве), которые сложились задолго до "прочтения" геномов. Аналогичные результаты получаются при сравнении практически любых генов в любых группах организмов. Каждый читатель может убедиться в этом самостоятельно, поскольку все прочтенные гены и программное обеспечение для их анализа находятся в свободном доступе.
Биохимическое единство жизни
Носителем наследственной информации во всех клетках являются молекулы ДНК, у всех известных организмов в основе размножения — репликация этой молекулы. В ДНК всех организмов используются 4 нуклеотида (аденин, гуанин, тимин, цитозин), хотя в природе встречаются не менее 102 различных нуклеотидов. Кроме того, в природе встречается 390 различных аминокислот, но белки всех организмов составляются из одного и того же набора, в котором всего 20 "основных" аминокислот и пара "дополнительных". При этом возможно 1.4 * 10^70 различных информационно эквивалентных генетических кодов, использующих те же самые кодоны и аминокислоты.
Как уже говорилось, если не учитывать эволюционное происхождение всех организмов от общего предка, то ничто не мешает каждому виду иметь собственный генетический код. Такое положение вещей было бы крайне выгодным, так как при этом исключалось бы преодоление вирусами межвидовых барьеров. Тем не менее, ничего подобного не наблюдается, и теория эволюции исключает такую возможность (см. выше).
Ученые, открывшие устройство генетического кода в 50-х и 60-х годах, в своих исследованиях активно использовали предположение, что код практически универсален. Эти ученые (Фрэнсис Крик, Сидней Бреннер, Георгий Гамов и другие) сделали это предположение исходя из версии об эволюционном происхождении от общего предка, не имея никаких прямых доказательств универсальности кода. Полагаясь на универсальность кода, Бреннер в 1957 году пришел к выводу о неперекрываемости кода (один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов) (Brenner, S. (1957) «On the impossibility of all overlapping triplet codes in information transfer from nucleic acid to proteins.»). Работа имела большое значение, так как до нее большинство исследователей считали код перекрывающимся.
В 1961 году, за пять лет до открытия устройства кода, Фрэнсис Крик сослался на работу Бреннера в статье «Общая природа генетического кода для белков». Основываясь на эволюционном предсказании об универсальности кода (в частности — на предположении о том, что у бактерий, табака и людей код устроен одинаково), Крик установил такие важнейшие свойства генетического кода, как триплетность (значащей единицей кода является сочетание трёх нуклеотидов — триплет, или кодон), неперекрываемость и наличие «стартовых» кодонов, с которых начинается чтение информации.
Единство генетического кода широко используется в биотехнологии. Например, бактериям встраивают ген человека, отвечающий за выработку инсулина, и бактерии начинают синтезировать человеческий гормон.
Также можно отметить крайне похожие пути метаболизма в клетках всех организмов. Так, гликолиз у всех эукариот и у большинства прокариот проходит за 10 одинаковых шагов, в одной и той же последовательности, с использованием одних и тех же десяти ферментов (при том что возможны тысячи различных, но термодинамически эквивалентных путей гликолиза). У всех изученных видов основным переносчиком энергии в клетке является аденозинтрифосфат (АТФ), хотя на эту роль могли бы претендовать сотни других молекул (см.: The fundamental unity of life из архива TalkOrigins.org).
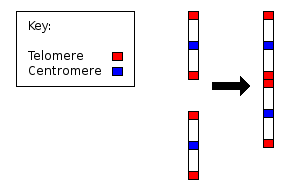
2-я хромосома человека
http://upload.wikimedia.org/wi...
После слияния двух хромосом остаются характерные следы: остатки теломер и рудимертарная центромера.
У всех человекообразных обезьян по 24 хромосомы, за исключением людей, у которых лишь 23 хромосомы. Человеческая 2-я хромосома является результатом слияния двух хромосом предков (Alec MacAndrew. Human Chromosome 2 is a fusion of two ancestral chromosomes).
Англ. ролик про 2-ую хромосому человека.
(Если ролика не видно на странице, его можно посмотреть здесь)
Доказательства слияния основываются на следующих фактах:
- Хромосома человека соответствует двум хромосомам обезьян. Ближайший человеческий родственник, бонобо, имеет практически идентичные находящимся 2-й хромосоме человека последовательности ДНК, но они расположены на двух отдельных хромосомах. То же самое верно и для более дальних родственников: гориллы и орангутана (см.: Comparison of the Human and Great Ape Chromosomes as Evidence for Common Ancestry)
- На хромосоме человека имеются рудиментарные центромеры. Обычно хромосома имеет только одну центромеру, но на длинном плече 2-й хромосомы наблюдаются остатки второй.
- Кроме того, на хромосоме человека имеются рудиментарные теломеры. Обычно теломеры находятся только на концах хромосомы, но последовательности нуклеотидов, характерные для теломер, наблюдаются ещё и в середине 2-й хромосомы.
2-я хромосома, таким образом, представляет собой убедительное доказательство эволюционного происхождения людей и других обезьян от общего предка.
Эндогенные ретровирусы
Эндогенные ретровирусы представляют собой следы древних вирусных инфекций в ДНК. Ретровирусы (такие как ВИЧ и Т-лимфотропный вирус человека, вызывающий лейкоз и лимфому) встраивают собственный геном в геном клеток зараженного организма. Обычно после этого клетка начинает продуцировать новые копии вируса. В этом процессе возможны сбои: копирование встроенной вирусной последовательности подавляется клеткой хозяина. Сама вирусная последовательность остается в структуре хромосомы. Если вирус встроился в геном стволовой половой клетки, то встроенную вирусную последовательность могут унаследовать потомки инфицированной особи. Ретровирусы встраиваются в геном случайным образом, вероятность независимой встройки одинаковых вирусов на одинаковые позиции у двух разных организмов пренебрежимо мала. А значит, встроенный геном одного и того же ретровируса может присутствовать у двух животных на одной и той же позиции в ДНК только в том случае, если эти животные произошли от общего предка.
Около 1 % человеческого генома занимают эндогенные ретровирусы, всего таких последовательностей в ДНК каждого человека около 30000 (Е. Д. Свердлов Retroviruses and primate evolution). Некоторые из этих ретровирусов встречаются только у человека. Другие последовательности встречаются только у шимпанзе и у человека, причем в одних и тех же позициях в геноме (тем самым подтверждается происхождение человека и шимпанзе от одного предка). Также есть последовательности, встречающиеся у горилл, шимпанзе и человека, у орангутанов, горилл, шимпанзе и человека, и так далее (см.: Endogenous retroviruses из архива TalkOrigins.org). Распределение эндогенных ретровирусов в точности соответствует филогенетическому дереву. См. также: Предки человека заимствовали полезные гены у вирусов
Также можно привести пример из семейства кошачьих. У малых кошек (точнее — у таких животных, как камышовый кот, европейская лесная кошка, степная кошка и домашняя кошка) найдена общая ретровирусная вставка. Ни у каких других хищников этот ретроген не обнаружен.
Ролик про эндогенные ретровирусы:
(если ролика не видно на странице, его можно посмотреть здесь)
Антиэволюционистам практически нечего возразить на совершенно убийственный для них "аргумент от эндогенных ретровирусов". Иногда они пытаются утверждать, что эндогенные ретровирусы якобы выполняют полезные функции в геноме многоклеточных организмов, и этим якобы объясняется их присутствие в одних и тех же местах генома у человека и шимпанзе и т.д. Но все эндогенные ретровирусы человека на самом деле неактивны, они давно утратили свою активность в результате мутаций. Другое дело, что в ходе эволюционного процесса, получившего название "молекулярное одомашнивание", некоторые отдельные фрагменты ретровирусных геномов (но не целые геномы) иногда "кооптируются" хозяином для выполнения каких-то полезных функций (см. в заметках: Предки человека заимствовали полезные гены у вирусов; Наездники подавляют иммунную защиту своих жертв при помощи прирученных вирусов). Большинство встроенных ретровирусных геномов, однако, совершенно бесполезны для хозяина, да и "молекулярное одомашнивание" никоим образом не может объяснить факт их присутствия в одних и тех же местах в геномах разных видов.
Псевдогены
Псевдогены - это неработающие, "молчащие" гены, которые возникают в результате мутаций, выводящих нормальные "рабочие" гены из строя (существуют и другие, более редкие пути возникновения псевдогенов). Псевдогены представляют собой настоящие "генетические рудименты". Если мутация выведет из строя ген, полезный для организма, она почти наверняка будет отсеяна отбором. Однако некоторые гены, в прошлом полезные, могут стать ненужными, например, из-за смены образа жизни. Мутация, выводящая из строя такой ген, не отсеивается отбором и может закрепиться в популяции. Псевдогены могут долго сохраняться в геноме в качестве ненужного "балласта". Мутации, которые в дальнейшем будут происходить в псевдогене, безразличны для выживания организма, и поэтому такие мутации свободно накапливаются и в конце концов могут изменить псевдоген до неузнаваемости. Однако на это уходит обычно много времени (десятки или даже сотни миллионов лет). Поэтому в геномах большинства организмов, включая человека, псевдогены на той или иной стадии мутационной деградации присутствуют в больших количествах. Псевдогены представляют собой своеобразную "историческую хронику", рассказывающую об образе жизни и адаптациях далеких предков изучаемого организма.
Например, в геноме человека в псевдогены превратились многие гены обонятельных рецепторов. Это и понятно, поскольку обоняние не имело существенного значения для выживания людей в историческое время, и, по-видимому, в доисторическое тоже (подробнее см. в заметке: Обоняние и цветное зрение в эволюции млекопитающих развивались в противофазе).
Ярким доказательством эволюции является присутствие одинаковых псевдогенов в одних и тех же местах генома у видов, произошедших сравнительно недавно от общего предка. Так, у человека есть псевдоген GULO, который представляет собой "сломанный" ген фермента глюконо-лактон-оксидазы. Этот фермент необходим для синтеза аскорбиновой кислоты. У других приматов обнаружен точно такой же псевдоген, причем мутационная "поломка", нарушившая работу гена, у него такая же, как и в человеческом псевдогене. Причины очевидны: в связи с переходом предков современных приматов к питанию растительной пищей, богатой витамином C, данный ген перестал быть необходимым для выживания. Мутация, испортившая ген, не была отсеяна отбором, закрепилась и была унаследована обезьянами и человеком. У других млекопитающих (например, у крысы) GULO является не псевдогеном, а работающим геном, и поэтому крысам не нужно получать витамин C с пищей: они синтезируют его сами. В других группах млекопитающих, которые независимо от приматов перешли к питанию пищей, богатой витамином С, тоже произошла псевдогенизация гена GULO, но мутации, выведшие ген из строя, у них были другие (пример - морские свинки).
Еще один пример: у млекопитающих есть три гена, которые у птиц и рептилий отвечают за производство белка вителлогенина, который входит в состав желтка в яйце. Почти у всех млекопитающих эти три гена - "мертвые", псевдогенизированные. Только яйцекладущие однопроходные звери (утконос, ехидна) синтезируют вителлогенин. У однопроходных из трех генов вителлогенина "мертвы" только два, а третий сохранил функциональность. Между прочим, хотя у плацентарных мелкопитающих желток не образуется, в ходе эмбриогенеза развивается рудиментарный желточный мешок (наполненный жидкостью), присоединенный к кишечнику зародыша. На втором месяце беременности у человека желточный мешок отделяется от эмбриона.
--------------------------------------------------------------------------------------------------------------------------
См. также: Ф. Коллинз. "Расшифровка божественных чертежей" (Глава из книги Фрэнсиса Коллинза "Доказательство Бога. Аргументы ученого". 2008). Полный текст книги в формате djvu. Ф.Коллинз - крупный американский генетик, руководитель проекта "Геном человека". В книге "Доказательство Бога" он приводит генетические аргументы в пользу эволюции и обосновывает свою точку зрения о том, что современная эволюционная биология, как и наука в целом, не противоречат христианству.





{kind=link}
Оценили 2 человека
4 кармы