

Автор материала: Василий Томсинский
Источник: http://evolution.powernet.ru/p...
Примечание: материал довольно объёмный, будет состоять из нескольких частей, поэтому, пожалуйста, наберитесь терпения, ибо оно того стоит (при условии, что вам интересна эта тематика). Пунктуация, орфография, ссылки и прочее - авторские. Я лишь делюсь информацией.
Введение.
Существование переходных форм – одно из самых захватывающих (если не самое захватывающее) свидетельств в пользу эволюции, в пользу вывода о том, что все живое связано одной общей родословной. Другие свидетельства вряд ли могут на равных тягаться с переходными формами. Атавизмы очень редки. Рудименты – части тела, существование которых не всегда очевидно, тем более у других животных. Гомология кажется просто сходством. В эмбрионах и компетентный биолог не всегда отыщет след эволюции, равно как и сами образцы. Молекулярное единство жизни – просто набор слов без точных знаний о внутриклеточных процессах. Понять значение генных деревьев трудно без знаний генетики, математики и систематики. Функции белков и их структуру в бытовых условиях не изучишь.
Иное дело переходные формы. Удивительно, что орел – это результат длительной модификации чего-то вроде ящерицы. Но вот окаменевшие кости животного, которое находится на промежуточной стадии этой модификации. Наша окаменелость – своего рода фотография процесса, который у нас нет никаких шансов наблюдать непосредственно. Преобразование ящерицы в птицу происходило более 200-80 миллионов лет назад. В то время как человечество изобрело письменность всего несколько тысяч лет назад, да и сам наш вид существует всего 200,000 лет. И тем не менее мы достаточно удачливы, чтобы иметь на руках реальную часть того эволюционного процесса. Как если бы мы могли отправиться в прошлое и поймать представителя промежуточной эволюционной группы. Мы можем изучить окаменелость по фотографиям, мы можем посмотреть на саму окаменелость через музейное стекло, мы можем сделать костям рентген, мы можем создать реконструкцию скелета, и даже можем попытаться реконструировать тело.
Переходные формы также не оставляют никаких сомнений насчет невозможности эволюции. Критики эволюционной теории говорят, что превращение ящерицы в птицу немыслимо, потому что оно потребовало бы кардинальной перестройки всех систем органов, появления сложнейшего механизма полета, возникновения удивительных инстинктов, что кажется невозможным. Однако переходные формы показывают, что это фактически произошло. Знаем мы, как это в точности случилось, или нет; кажется ли нам это невероятным или нет; можем ли мы в это поверить или нет – не имеет никакого значения. Эволюция произошла, как не крути.
Я подчеркнул ценность переходных форм лишь для того, чтобы теперь показать, что их иллюстративность слишком переоценивается. Они убедительны для специалистов. Для обычных людей существование переходных форм в большей мере вопрос веры, поскольку они не в состоянии оценить, почему те являются переходными. Это очень важное обстоятельство, которое популяризаторы науки почти всегда игнорируют. Есть несколько проблем с пониманием переходных форм.
Во-первых, по окаменелостям исследовать эволюцию беспозвоночных или растений существенно сложнее, чем эволюцию позвоночных. А небиологи главным образом не в состоянии сравнить строение беспозвоночных или растений. Следовательно, приходится ограничиться только позвоночными животными. Но в результате возникают подозрения такого рода:
"Если эволюционисты вообще ссылаются на свидетельства окаменелостей, то лишь для некоторых групп позвоночных организмов, так как рыбы, земноводные, пресмыкающиеся и млекопитающие образуют определенную последовательность в отложениях".
Уильям Гладсхауэр. "Как возник наш мир" [124].
Во-вторых, переходные формы представлены окаменевшими останками. Находки полностью сохранившихся скелетов - редкость. Отдельные кости сохраняются частично и могут накрывать друг друга. Требуется специальная подготовка и скрупулезная работа для анализа экземпляра. Поэтому фотографии окаменелостей не могут сказать почти ничего обычному человеку с улицы. Скорее всего, они произведут на него удручающее впечатление. Можно продемонстрировать реконструкцию скелета и надеяться, что читатель разберется в его строении сам. Другой вариант (самый распространенный) – нарисовать переходные формы такими, какими они могли бы быть при жизни. Правда, фантастичность таких картин довольно очевидна, и вряд ли они что-то демонстрируют. Вся очевидность их промежуточного эволюционного положения оказывается скрытой под нарисованными покровами.
В-третьих, – и это главная проблема, – неспециалист не знает анатомию и просто не в состоянии заметить, что перед ним переходная форма. Требуется несколько секунд, чтобы показать человеку окаменелость или ее реконструкцию, и затем нужно потратить не меньше часа на лекцию об анатомии скелета разных групп позвоночных животных. Причем необходимо, чтобы слушатель запомнил материал и мог в нем самостоятельно ориентироваться. Кто без такой лекции сможет сказать, сколько позвонков должно быть в шейном отделе рептилий и птиц, какая форма позвонков должна быть у каждой группы?
Именно поэтому Кен Миллер, блестящий полемист, решив доказать существование переходных форм, смог лишь показать несколько иллюстраций внешне похожих скелетов и описать работу палеонтологов [53]. Антрополог Джим Фоли, обсуждая ископаемых гоминид, сконцентрировался на двуногом передвижении и размере мозга. Палеонтолог Дональд Протеро, описывая многочисленные переходные формы, обычно ограничивается указанием двух-трех особенностей их анатомии. (Однако, на мой взгляд, он преуспел больше других в популяризации палеонтологии и эволюции, и я настоятельно рекомендую его замечательные книги всем, кто интересуется данной темой) [64, 65, 66].
Археоптерикс – пример переходной формы №1. "Берлинский экземпляр археоптерикса – самая важная находка палеонтологии", – сказал как-то палеонтолог Джон Остром. Но все же популярные издания описывают археоптерикса неизменно однобоко. Из птичьих особенностей называются оперение, а из рептилийных – когти, зубы и длинный хвост. Множество других анатомических черт, более важных, остаются неупомянутыми. Здесь нечему удивляться. Подробный анатомический обзор сделает описание нечитабельным. Но в то же время скупость популярных описаний производит ложное впечатление. Люди думают, что больше об археоптериксе и сказать-то нечего, и приходят к заключению, что палеонтологи делают поспешные выводы на основе столь скудных данных.
Подводя итог, можно сказать: не важно, существуют переходные формы или нет, так как даже если они существуют, вы не сможете это продемонстрировать без длительного курса палеонтологии и анатомии. И популярные описания эволюции, называя некоторых вымерших животных и объявляя их переходными, не достигают поставленной цели. Мало того, они еще и создают искаженное впечатление об эволюционной биологии.
В этой статье я попытаюсь закрыть пробел между популярными книгами и профессиональной палеонтологией, на материале скелетной анатомии рептилий и птиц.
Часть 1. Переходы и переходные формы.
Археоптерикс и происхождение птиц.
Существование переходных форм является необходимым следствием теории единой общей родословной всех форм жизни, которая составляет основу эволюционной биологии. В соответствии с этой теорией, какие бы формы жизни мы не взяли в качестве примера (при условии, что одна форма не является прародителем второй), они будут потомками общего предка, который существовал в некоторый момент в прошлом. Степень родства и соответственно древность общего предка может значительно изменяться. Так оценено, что человек разумный и шимпанзе связаны весьма близким родством. Общий предок этих видов жил примерно 6 миллионов лет назад. Человек разумный и неандерталец связаны еще более близким родством. Их прародитель существовал, согласно самой точной оценке, около 700,000 лет назад. По контрасту человек и сизый голубь имеют гораздо более дальнее родство. Их генеалогические линии разошлись около 300 миллионов лет назад. Графическим представлением теории единой общей родословной является всем известное эволюционное древо или в более современном варианте - кладограмма.
Теория единой общей родословной предполагает, что происхождение новых биологических видов, родов, семейств, отрядов и т. д. происходит путем преобразования (эволюции) биологии (в том числе строения) предка, которому они обязаны своим появлением. Например, птицы возникли в результате преобразования биологии наследственной для них группы рептилий. А значит, должны существовать формы, которые находились на различных стадиях этого преобразования. Однако среди ныне живущих организмов таких найти проблематично. В этом нет ничего удивительного. Мы знаем, что многие группы организмов существуют на Земле очень давно. Например, птицы живут на планете не менее 150 миллионов лет, с Юрского периода, если вы предпочитаете относительную хронологию. Стало быть, преобразование строения некой группы древних рептилий, которое в конечном итоге привело к появлению птиц, происходило в те далекие времена.
Учитывая сказанное, теперь мы можем определить, что же такое "переходные формы". Переходные формы - это такие формы жизни, которые демонстрируют преобразование одного уникального типа строения, характерного только для одной группы организмов (родительской), в другой уникальный тип, характерный для другой группы (потомковой). Таким образом, переходная форма должна обладать комплексом уникальных черт строения одновременно и предковой, и потомковой групп. Если теория единой общей родословной верна, переходные формы обязательно должны были существовать. И мы можем попытаться отыскать их в окаменелостях.
В этом разделе я опишу археоптерикса и покажу, что он занимает промежуточное положение между мезозойскими рептилиями и птицами, причем анатомически он даже ближе к рептилиям, чем к птицам. Но следует сразу оговориться, что это будет лишь частичное и упрощенное описание. Я буду использовать только наиболее простые признаки. Палеонтологи в своих исследованиях стараются учесть любую мелочь, поэтому их работа обычно включает длинные перечни и простых, и тончайших различий в анатомии. Однако включение подробностей такого рода здесь сделает текст нечитабельным. И хотя наиболее сложные различия останутся за скобками, следует помнить, что они есть, и все последующее описание не дает полной картины.
Для наглядности я предлагаю читателю преобразовать его собственный скелет в скелет археоптерикса. Мы можем это себе позволить благодаря гомологии. Широко распространено заблуждение, что гомология - это сходство в строении. В действительности гомология имеет более существенное основание. В гомологичных структурах составляющие их элементы расположены по типовому образцу и имеют типичную структуру.
Чтобы понять гомологию, стоит вспомнить Дарвина. Биологи задолго до Дарвина отмечали, что у разных и несхожих организмов образец строения органов и их расположения совпадает. Но почему это так, никто удовлетворительно объяснить не мог. Распространенность гомологии подталкивала разных исследователей к мысли о том, что в некоторых случаях она является результатом происхождения от общего предка. Однако именно Дарвин придал этому аргументу современную форму. Суть идеи в том, что новые биологические черты организмов возникают путем постепенного преобразования старых. С этой точки зрения, например, организм человека – это модифицированный организм рыбы. Даже когда появляются новые элементы, например, новые кости, они возникают из преобразования старых структур. Именно так возникает гомология. Скелет всех четвероногих животных – это модификация базового образца строения древних амфибий и рыб. То же верно для прочих систем организма – нервной, кровеносной, выделительной и т. д. И чем ближе родство, тем больше гомологическое сходство.
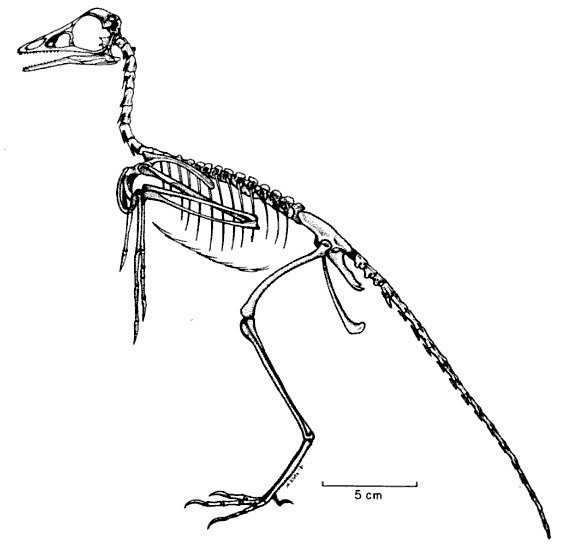
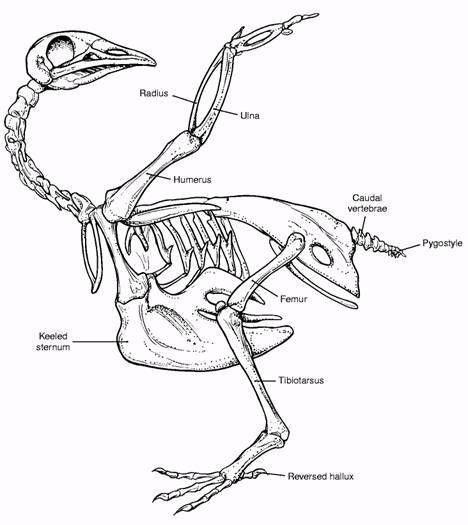
Здесь я поместил для сравнения рисунки скелетов археоптерикса и голубя. Если вы вернетесь к ним после сравнения анатомии археоптерикса, его ближайших родственников и птиц, уверен: схемы покажутся более информативными. Можно также рассмотреть интерактивную схему скелета голубя, а также подборку трехмерных моделей скелетов разных птиц.
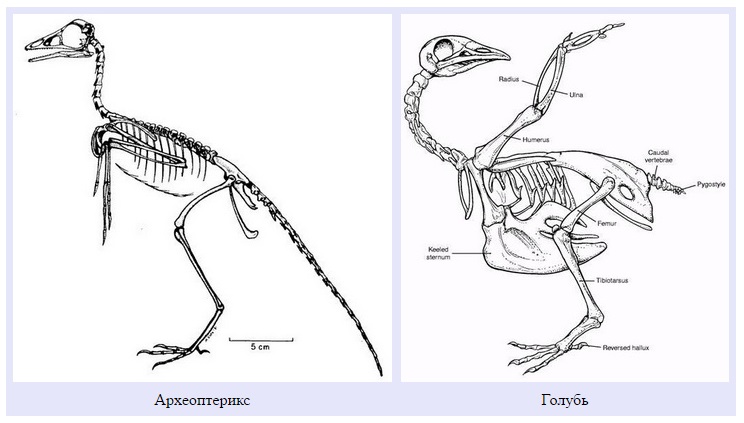
Итак, читатель, сделаем из Вас гордую мезозойскую птицу.
Чтобы стать ею для начала Вам придется перестать быть млекопитающим. У Вас три косточки в среднем ухе и одна кость в нижней челюсти, а нужно, чтобы в нижней челюсти было несколько костей и одна слуховая в среднем ухе. Форма черепа совершенно неудовлетворительна, он слишком овальный. Нельзя стать археоптериксом без грубо треугольного, вытянутого черепа. Но главное все же – проделать перед глазницей еще парочку отверстий. Особенно гордость всех архозавров – предглазничное окно и ближе к ноздрям скромное предглазничное отверстие. Чуть выше добавим утерянную предлобную кость. За глазницей тоже нужно поработать. Туда придется приладить еще одну потерю – заглазничную кость.

Небные кости явно не в порядке. Как минимум нужно добавить то, что называется наружной крыловидной костью.
У Вас есть зубы – это хорошо, но они слишком разнообразны, целых четыре типа. Археоптериксу такие излишества ни к чему. Отныне у Вас будут однообразные острые конические зубы. А между зубами расположатся межзубные пластины – маленькие костистые выросты на челюстях. И хотя Вам теперь будет сложнее пережевывать пищу, зато зубы могут сменять друг друга неограниченное число раз – на месте старого зуба неизменно будет вырастать новый.
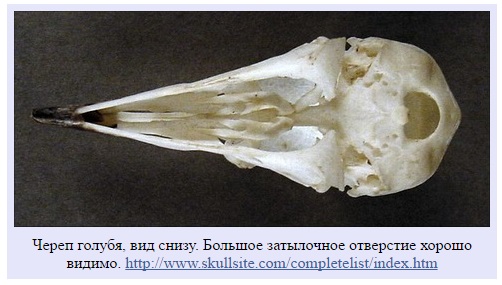
Далее. Голова соединяется с шеей не в том месте. Чтобы получить право называться археоптериксом, придется оторвать голову, поместить большое затылочное отверстие сзади черепа, а не на дно, и снова соединить голову с шеей.
Далее. Суставные поверхности ваших шейных позвонков плоские. Это эстетика млекопитающих, в мезозое для нее еще не пришло время – придется поменять на двояковогнутые.
Далее. В крестцовом отделе позвоночника у Вас 5 позвонков. Именно столько и надо, чтобы быть археоптериксом. Но если Вам уже исполнилось 25 лет, то все ваши пять позвонков срослись в единое тело. Так не пойдет. Придется позвонки разъединить.
Далее. Кости таза. С такими лобковыми костями вам ничего не светит в мире археоптериксов. У вас они срастаются, но и только-то. Придется их вытянуть, отдаленные концы снабдить расширением в форме ботинка и уже тогда срастить отдаленные концы.
Далее. Смущает отсутствие хвоста. У вас есть несколько деградировавших позвонков, сросшихся в единое образование (копчик). Такого археоптерикса никто уважать не будет. Хвост должен быть длинным из двадцати цельных позвонков. Чем ближе к кончику хвоста, тем более вытянутыми должны становиться тела позвонков.
Далее. Передние конечности. С точки зрения археоптерикса, совершенно не ясно, зачем Вам столько пальцев. Оптимальное число – 3. Так что отбрасываем безымянный и мизинец. Придется также пересчитать количество фаланг пальцев. У вас по три фаланги в каждом из оставшихся пальцев. Это никуда не годится. В большом пальце две фаланге, в указательном – три, это правильно, а вот к безымянному одну нужно прибавить. Еще мы добавим вам изогнутые когти. Можете считать это компенсацией. Кроме того, ваше запястье довольно однообразно. Вам просто необходимо срастить первую и вторую кости запястья в единое образование и придать ему полулунную форму. Эта штука, называемая полулунной костью запястья, позволяет сворачивать кисть по-птичьи.
И не забудем про лопатки. У вас к лопатке прирастает еще одна кость – коракоид или вороновидная кость, образующая с лопаткой единое тело. Археоптериксу полагается иметь эту кость отдельной, самостоятельной и хорошо развитой, грубо квадратной формы. Причем этот коракоид должен иметь выступающий вперед отросток. Что же до лопаток, то они должны расширяться в отдаленном от плечевого сустава конце.
Далее. Ваши ключицы нуждаются в немедленном исправлении. Необходимо срастить их концы в единую кость – вилочку. И пусть грудина разделяется на отдельные части.
Далее. От бывшего млекопитающего мало что осталось. Не будем придираться к ребрам. Но добавим вам брюшные ребра – типичный признак пресмыкающихся. Теперь даже искушенный глаз не узнает в вас заурядного млекопитающего.
Далее. Ваши задние конечности практически не нуждаются в изменениях. Хотя одного пальца придется лишиться. А фаланги второго пальца придется растянуть так, чтобы этот палец стал самым длинным и загнутым кверху. Затем мы приделаем к нему хищный серповидный коготь. Кроме того, таранная кость должна иметь отличительный восходящий отросток.
Почти все. Осталось сделать несколько общих изменений.
Во-первых, у вас слишком тяжелые кости. Они явно нуждаются в пустотах, заполненных воздухом. От этого скелет станет только легче.
Во-вторых, вам пригодится оперение. Асимметричные перья на передних конечностях, пучок длинных перьев на конце хвоста и контурные перья тела.
Теперь вы очень похожи на археоптерикса, но в целом выглядите вы чудовищно. Чтобы стать археоптериксом, нужно еще уменьшиться в размерах. 50 сантиметров от кончика морды до кончика хвоста, 25 сантиметров в высоту. Это самое большее, что вы можете себе позволить.
Теперь, когда у нас есть археоптерикс, мы можем начать модифицировать его в птицу. Попутно я буду сравнивать археоптерикса с его ближайшими родственниками – дейнонихозаврами – группой звероногих динозавров. Вы сможете сами оценить, является ли строение археоптерикса в большей мере птичьим или рептилийным. Начнем быструю эволюцию, на которую у природы ушло около 80 миллионов лет.
Зубы и межзубные пластины. Все современные птицы являются беззубыми, межзубные пластины также отсутствуют. Зубы закладываются в эмбрионах птиц, но на более поздней стадии рассасываются. С помощью манипуляций с эмбриональными тканями ученые даже смогли получить цыплят с зубами. Эти данные показывают, что генетические схемы, ответственные за развитие зубов, все еще присутствуют в геноме птиц. У ближайших родственников археоптерикса зубы были. Как мы теперь знаем, потеря зубов у птиц происходила в разных эволюционных линиях независимо и в разное время. Некоторые птицы утратили зубы в начале мелового периода. Последние зубатые птицы вымерли 60 миллионов лет спустя. Роговой чехол, покрывающий челюсти – клюв – очевидная птичья особенность. Но ее придется добавить, так как археоптерикс и дейнонихозавры не имели такого чехла.
Череп нуждается во множественных модификациях. В отличие от археоптерикса и дейнонихозавров у птиц утрачены предлобная, заднеглазничная и наружная крыловидная кости, но предглазничное окно остается.
Характерная особенность черепа птиц - подвижность и верхней, и нижней челюсти, что контрастирует с типичным состоянием у большинства четвероногих животных (включая человека), у которых подвижна только нижняя челюсть. Механику челюстей археоптерикса установить непросто, но анатомия верхних челюстей несовместима с тем типом подвижности, который типичен для современных птиц.
Большое затылочное отверстие на том же месте, что у дейнонихозавров, но не птиц. У птиц оно смещено на дно черепа.
Шейные позвонки археоптерикса с их двояковогнутыми суставными поверхностями такие же, как у ближайших родственников. По контрасту шейные позвонки птиц имеют седловидные суставные поверхности. Это предмет особой гордости птиц, они уникальны. Ни у каких других животных нет таких позвонков, и никогда не было. Исключение составляют цератозавры – группа триасовых звероногих динозавров. Однако у них эта особенность была не столь выражена, как у птиц. Благодаря этой уникальной адаптации птицы могут поворачивать голову на 180 градусов, а некоторые на 270.

У археоптерикса и дейнонихозавров крестцовый отдел позвоночника насчитывает то же количество свободных позвонков. У птиц этот отдел сильно изменен. Число позвонков в нем увеличено (колеблется от 11 до 23), позвонки срастаются между собой в так называемый сложный крестец. Также у птиц тазовые кости срастаются между собой. Напротив, лобковые кости у птиц не срастаются, а их отдаленные концы лишены расширения в виде ботинка.

Хвостовой отдел у птиц редуцирован – хвостовые позвонки упрощены, срастаются между собой и в целом хвостовой отдел сливается со сложным крестцом. Заканчивается хвост пластинкой – пигостиль – это важнейшая адаптация. Пигостиль служит для прикрепления рулевых перьев, таким образом, он обеспечивает совершенство птичьего полета. Пресловутый длинный хвост археоптерикса – полная противоположность состоянию хвостового отдела у птиц. В популярной литературе он систематически упоминается неправильно. Дело не в длине. Длина сама по себе ничего не значит. В природе избыток хвостов любой длины. Но у археоптерикса хвостовые позвонки не срастаются и не уменьшаются, чем ближе к кончику хвоста, тем более вытянутыми, цилиндрическими становятся тела позвонков. Выглядит это так, как будто археоптерикс украл свой хвост у дейнонихозавров.
По строению передних конечностей дейнонихозавры приняли бы археоптерикса за своего. Те же три пальца, с такими же когтями, то же число фаланг. Но у птиц передняя конечность устроена иначе. Археоптерикс, дейнонихозавры и птицы, все трехпалы. Четвероногие животные (кроме самых ранних и самых древних) первично имеют пять пальцев. Иное число пальцев в разных группах четвероногих связано с утратой того или иного их числа. В эмбрионах птиц все еще можно отыскать след их происхождения от древних пятипалых предков. В течение эмбрионального развития у птиц закладываются пять пальцев, но два в итоге рассасываются. Три остающихся пальца редуцированы, присутствует лишь несколько видоизмененных фаланг, служащих для закрепления маховых перьев. Число фаланг у птиц 1-2-1 в противоположность 2-3-4 у археоптерикса и дейнонихозавров.
Кроме того, у птиц кости пястья и передний ряд костей запястья срастаются в единую пястнозапястную кость. Если вам не нравится такое название, вы можете называть ее пряжкой. Это тоже будет правильно. У дейнонихозавров все кости запястья и пястья свободны, но об археоптериксе такого не скажешь. У него кости пясти не срастаются, но сильно сдавлены, таким образом, пряжка еще не сформирована, но начало процессу положено.
Полулунная кость запястья, которая есть у археоптерикса - это, скорее, птичий признак, но птицы разделяют эту особенность не только с археоптериксом, но и с дейнонихозаврами, у которых она является самостоятельным образованием. У птиц полулунная кость входит в состав пряжки. Она позволяет сворачивать кисть так, что та оказывается почти параллельна предплечью.

Подобно своим динозавровым родственникам археоптерикс не имел надкаракоидного сухожилия. Эта важная адаптация позволяет птицам поднять крыло выше спины, тем самым птица может сделать мощный взмах, необходимый для машущего полета. Не ясно, был ли археоптерикс способен к машущему полету. С одной стороны, отсутствие надкоракоидного сухожилия показывает, что поднять крыло выше спины он не мог, а с другой – какая-то примитивная форма машущего полета, не свойственная современным птицам, у него все же могла быть.
Лопатка археоптерикса, как и у дейнонихозавров, в отдаленном конце имеет расширение. У птиц такого расширения нет. Суставная ямка лопатки у археоптерикса занимает промежуточное боковое положение. У птиц она смещена в спинном направлении. А связанный с лопаткой коракоид по образцу птиц имеет выступающий отросток. У дейнонихозавров этой особенности нет. В соответствующем положении на коракоиде у них присутствует бугристость, которая рассматривается палеонтологами как предшественник выступающего отростка. Сам корокоид археоптерикса устроен по образцу дейнонихозавров, а не птиц. Он плоский и имеет грубо квадратную форму. У птиц коракоид вытянут и принимает форму распорки.
Птицы также отличаются и от археоптерикса, и от дейнонихозавров наличием киля – выроста грудины, служащего для прикрепления летательной мускулатуры.
Ключицы, сросшиеся в вилочку – еще один признак, который роднит птиц, археоптерикса и дейнонихозавров. А ребра разделяют группы. Брюшные ребра есть у археоптерикса, его родственников и множества рептилий, у птиц их нет. Грудные ребра птиц скрепляются между собой выростами – крючковатыми отростками. Их нет у археоптерикса, хотя они есть у некоторых дейнонихозавров.

По строению нижних конечностей археоптерикс с его удлиненным вторым пальцем, украшенным хищным серповидным когтем, присоединяется к компании дейнонихозавров. В конце концов, именно из-за этого когтя они получили свое имя – "ужасный коготь". Археоптерикс также разделяет с дейнонихозаврами строение голени и стопы. Стопа и голень птиц перестроены. Прежде всего, у птиц редуцирована малая берцовая кость. Она уменьшилась до тонкой щепы и приросла к большой берцовой. У археоптерикса малая берцовая кость полностью развита. Ближе к стопе – у птиц пяточная и таранная кости срастаются друг с другом и с большой берцовой. Археоптерикс эту характеристику также не разделяет. Более того, таранная кость археоптерикса снабжена развитым восходящим отростком. Птицы эту характеристику утратили. Она обнаруживается в течении эмбрионального развития, но исчезает на более поздних стадиях. Здесь анатомия археоптерикса снова соответствует типу строения ближайших родственников, но не современных птиц. Птицы обзавелись цевкой – костью, образованной срастанием части костей плюсны и предплюсны. У археоптерикса цевки нет, но, как и в случае с пряжкой, соответствующие кости сдавлены. Таким образом, цевка находится в стадии формирования.

Оперение – то, что отличает птиц и археоптерикса от всех ныне живущих животных, но не от дейнонихозавров. Последние также обладали оперением. Причем некоторые имели асимметричные перья (асимметрия пера – необходимое условие для полета).
Как легко заметить, у археоптерикса нет почти ничего специфически птичьего. Его анатомия в гораздо большей степени рептилийная, чем птичья. В скелете археоптерикса сочетаются одновременно черты дейнонихозавров и птиц, поэтому он представляет собой мост между дейнонихозаврами и более поздними птицами, которые в свою очередь промежуточны по сравнению с современными птицами. Если бы мы сделали этот вывод 40 лет назад, могли бы претендовать на открытие. К сожалению, нас опередил палеонтолог Джон Остром. Он предпринял скрупулезный обзор анатомии археоптерикса и всех потенциально родственных групп животных. В итоге Остром заключил, что археоптерикс по существу – маленький оперенный динозавр.
Следующая диаграмма показывает числовое соотношение рептилийных и птичьих особенностей анатомии у ранних птиц и наиболее близких к ним динозавров. Список сравниваемых особенностей см. в приложении 3.
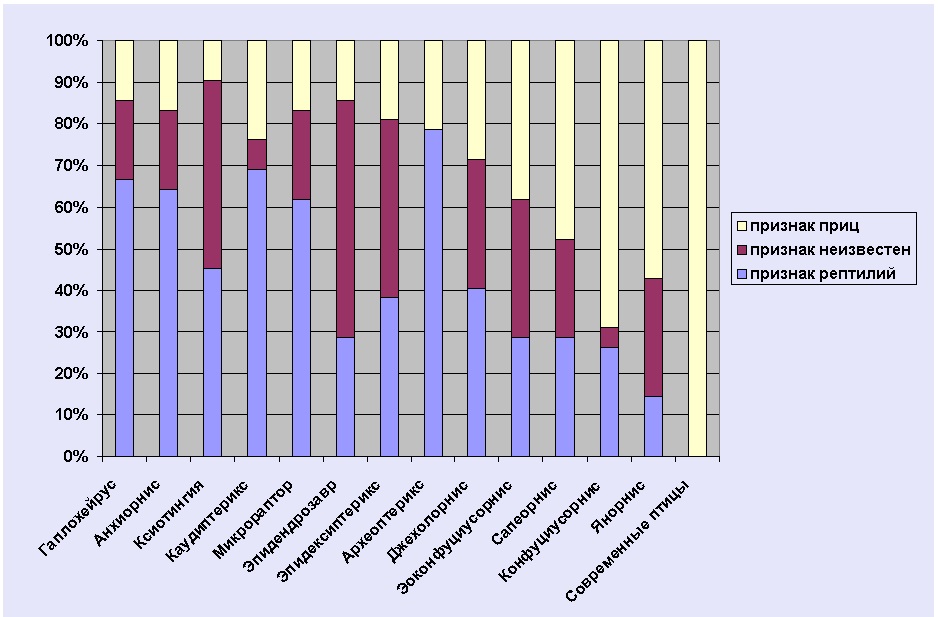
Я говорил, что представленное сравнение археоптерикса и его родственников является частичным и упрощенным. Для составления диаграммы я использовал 42 ключевых признака, наиболее простых и понятных. Палеонтологи обычно используют в своих исследованиях от 100 до 300 признаков. При этом они сравнивают значительно большее количество представителей и используют алгоритмы, основанные на теории вероятностей. Я надеюсь, что это даст некоторое представления об уровне упрощения, на котором нам приходится говорить об изучении переходных форм (см. также примечание a).
Как видно из диаграммы, археоптерикс действительно может быть описан как оперенный динозавр. Кроме того, он не одинок в своем роде. И если археоптерикс стоит ближе к динозаврам, чем к птицам, то другие формы, например Эоконфуциусорнис, находятся где-то на полпути.
Таким образом, единство в скелетной анатомии и существование ряда переходных форм указывают на происхождение птиц от одной из линий звероногих динозавров. Но кости не одиноки. Есть ряд других, несвязанных данных, подтверждающих этот вывод. Действительно, если птицы ведут свою родословную от общего предка с дейнонихозаврами, то мы должны обнаружить единство в аспектах биологии между птицами и близкими к ним звероногими. В то же время, биология птиц и звероногих должна контрастировать с биологией других групп животных. Все, что нам остается сделать, сравнить биологию птиц и звероногих в тех аспектах, которые нам известны.
Размер тела.
Способность к полету накладывает сильное ограничение на размер тела. Неслучайно птицы, утратившие способность к полету, могут приобретать очень большой размер тела (например, страусы и моа).
По контрасту, те дейнонихозавры, анатомия которых в прошлом служила эталоном для сравнения с птицами – дейноних и велоцираптор – крупные животные. Физически невозможно, чтобы животные такого размера могли освоить полет. Поэтому долгое время приходилось предполагать, что более близкие родственники птиц были существенно меньше в размере. Предположение было подтверждено с открытием микрораптора, который был описан в 2000 году как "самый маленький из известных динозавров". Девять лет спустя был описан анхиорнис – дейнонихозавр еще меньшего размера. Фактически, анхиорнис даже меньше микрораптора. Анализ скелетной анатомии показывает, что анхиорнис и микрораптор стоят ближе других к птицам. На сегодняшний день они остаются самыми маленькими и легкими из всех динозавров [39, 80, 98, 99].

Мозг.
В данном случае обнаруживаются еще более детальные различия. Мозг современных птиц значительно отличается от мозга рептилий. Полушария головного мозга у птиц сильно увеличены. Также чрезвычайно увеличились мозжечок и зрительные доли. Что, конечно, связано со способностью к полету. По контрасту обонятельные луковицы сократились в размерах.

Мозг не окаменевает. Означает ли это, что мы никогда не сможем ничего узнать о мозге вымерших животных? Не обязательно. Головной мозг окружен твердой мозговой оболочкой, которая отражает строение мозга. И если череп хорошо сохранен, есть возможность создать (физический или предпочтительно компьютерный) слепок мозга.
Мозг большинства звероногих динозавров, чьи черепа позволяют реконструкцию, в целом устроен по типичному для археозавров образцу. Но эта картина начинает изменяться, когда мы обращаемся к ближайшим родственникам птиц. В отличие от других звероногих, их мозг ближе к птичьему образцу: относительный размер больших полушарий, мозжечка и зрительных долей увеличивается, однако, не достигает характерных для птиц соотношений. Обонятельные луковицы, напротив, остаются развитыми или даже увеличиваются. Сокращение этих частей мозга произошло позднее. Действительно, обонятельные луковицы археоптерикса все еще соответствуют рептилийному, а не птичьему образцу. В целом, мозг археоптерикса характеризуется промежуточным состоянием между рептилийным и птичьим, его <мозжечок и большие полушария увеличены, но не столь существенно, как у современных птиц [1, 31, 43, 45, 60].
Размножение.
Птицы, как и большинство рептилий, размножаются, откладывая яйца. Форма яиц отличается в различных группах. Яйца крокодилов овальные. У черепах – овальные или шарообразные, у большинства динозавров – шарообразные. Яйца птиц асимметричны – один полюс яйца шире другого. Яйца звероногих, близкородственных птицам, также асимметричны.
Еще более важная характеристика яиц – микроструктура скорлупы. Микроструктура птичьих яиц существенно отличается от микроструктуры рептилий, включая крокодилов, завроподов и птицетазовых динозавров. Однако она практически идентична у птиц и близкородственных звероногих!
Черепахи, змеи, крокодилы кладут яйца однократно. То же справедливо в отношении динозавров. Мы знаем это, поскольку яйца в кладках динозавров лежат беспорядочно единой группой. По контрасту птицы откладывают не все яйца разом, а последовательно через промежутки времени, что связано с ограничением в размерах тела. Изучая сохранившиеся кладки близких родственников дейнонихозавров и птиц, – овирапторидов – палеонтологи обратили внимание, что в их гнездах яйца располагаются не в общей массе, а объединены в пары. Такое пространственное расположение интерпретируется как признак того, что самки овирапторидов откладывали по два яйца через некоторое время. Гипотеза подтверждается находкой овирапторида только с двумя яйцами в полости тела.

Название овирапторидов восходит к описанию в 1924 году динозавра, останки которого были обнаружены поверх гнезда. Палеонтологи заключили, что это был хищник, разорявший чужие гнезда. Таким образом, динозавр получил имя овираптор – крадущий яйца. Однако открытие новых экземпляров, заставило пересмотреть вывод. Были описаны экземпляры овирапторидов, сидевших в центре гнезда в типично птичьей позе и накрывавших яйца передними конечностями. Выяснилось также, что подобное поведение было характерно и для некоторых дейнонихозавров. Неизвестно, с чем именно связано такое поведение. Одно очевидное объяснение связывает присутствие родителя с охраной кладки, другое – с буквальным высиживанием яиц. Независимо от того, какое из этих предположений правильное, данные показывают, что близкие родственники птиц заботились о своих кладках так же, как и современные птицы. Возможно, даже согревали яйца теплом своего тела. Такое предположение не кажется невероятным, если вспомнить, что близкие родственники птиц (овирапториды здесь примыкают к дейнонихозаврам) имели оперение [5, 8, 35, 38, 42, 44, 103, 104].
Перья.
Поскольку в оперении археоптерикса присутствуют перья с типично птичьей структурой, было ясно, что археоптерикс мог унаследовать их от предков (см. приложение 1). Предположение подтвердилось только на рубеже XX и XXI веков, когда впервые были описаны овирапториды с сохраненными перьями. На сегодняшний день перья известны в трех наиболее близких к птицам семействах звероногих и не в каких других. Описаны звероногие с симметричными и асимметричными перьями Ннапомню, асимметрия – необходимое аэродиномическое условие для полета. В общих чертах оперение близких родственников птиц соответствует птичьему: пучок длинных перьев на кончике хвоста (рулевые перья у птиц), длинные и крупные перья на передних конечностях (маховые перья у птиц) и контурные перья тела [41, 56, 78, 81, 93, 94, 118].
Поведение.
В 2005 году палеонтологи представили миру экземпляр хорошо сохранившегося дейнонихозавра, у которого голова была повернута назад, а морда частично накрыта передней конечностью. Аналогичное положение тело можно наблюдать у современных птиц во время сна. Голова у птиц, как предплечья и голени у человека, именно та часть тела, которая обеспечивает наибольшую потерю тепла и соответственно охлаждение тела. Поэтому птицы типично во время сна накрывают голову крылом, что уменьшает теплопотерю. Предположительно, упомянутый динозавр спал в момент внезапной смерти [101].

Перечисленные здесь особенности указывают на единство биологии одной ветви звероногих динозавров и птиц. Однако обратите внимание, что проведенное сопоставление не имеет отношения к переходности или непереходности археоптерикса и прочих форм, о которых шла речь выше. Переходный статус зависит от анатомии. Сравнение биологии птиц и родственных звероногих преследует другие цели. Оно демонстрирует:
Во-первых, интегральный характер изучения эволюции, предполагающий привлечение информации из разных областей биологии.
Во-вторых, как на основе выделения переходных форм можно выдвигать гипотезы, в поддержку которых постепенно накапливаются данные.
Тираннозавр наполовину.
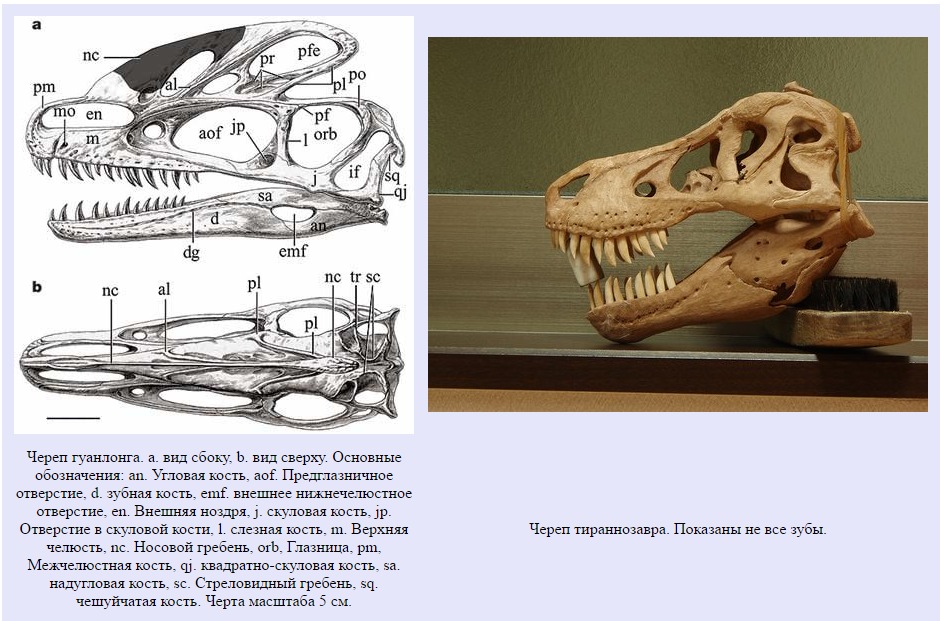
"Точные предки тираннозавров невыяснены". Палеонтолог Роберт Кэррол в 1989 году [128].
Чтобы искать предков какой-либо группы животных, сначала нужно установить диагностические черты, присущие только ее представителям. Тираннозавриды – типичные хищные звероногие динозавры. Это часто крупные животные, передвигавшиеся на двух задних конечностях. Их зубы крупные, зазубренные и загнутые назад. Эти зубы не оставляют сомнений на счет кулинарных пристрастий. Они имели большие головы и большие хвосты, служившие противовесом для головы и туловища. Но чем они отличались от других звероногих и своих ближайших родственников? Правильная классификация окаменелостей – сложная задача, и вы сейчас в этом убедитесь, поскольку нам придется углубиться в еще более сложные тонкости скелетной анатомии.
У тираннозаврид:
* Носовые кости срастались друг с другом.
* Теменные кости также срастались, образуя стреловидный гребень.
* Зубы, связанные с межчелюстной костью обладают U-образной формой.
* Сама межчелюстная кость имеет большое боковое отверстие.
* Основная клиновидная кость имеет большую подмыщелковую полость.
* Надугловая кость снабжена выраженным прямым гребнем.
* Досочленовный отросток укорочен.
* Скуловая кость частично пневматизирована и имеет окно.
* Чешуйчатая кость в задней части имеет большое отверстие.
* Отверстие в задней части нижней челюсти значительно уменьшено.
* Верхняя затылочная кость не формирует большое затылочное отверстие.
* Поперечные отростки шейных позвонков вытянуты. Они служат для прикрепления мощных мышц, удерживающих большую голову.
* Передние конечности чрезвычайно маленькие относительно задних. Функция столь маленьких конечностей не выявлена до сих пор.
* Конец лучевой кости сглажен.
* Присутствуют только два пальца.
* Второй палец передней конечности существенно больше.
* Лобковая кость заканчивается очень большим расширением в форме ботинка (около 50% длинны всей кости).
* Подвздошная кость примерно равна в длине бедру.
* Третья кость в плюсне сжата второй и четвертой. Такая структура называется арктометатарсус. Арктометатарсус – адаптация к бегу.

Если вы обескуражены этим панегириком анатомии, утешайтесь тем, что это еще упрощенный список.
Теперь, когда мы определили отличительные особенности тираннозаврид, можно искать переходные формы к ним. Но самое интересное, что мы можем частично предсказать, на что будет похожа такая форма. Если мы знаем ближайших родственников тираннозавров, значит, сравнивая анатомию всех связанных групп, можно вывести черты, которые разделили их предковые формы. Если мы правильно определили родство, то переоценка уже описанных окаменелостей и/или обнаружение новых подтвердят гипотезу. В противном случае они опровергнут построенную схему эволюционных отношений. В сущности, именно так выдвигаются и проверяются эволюционные гипотезы.
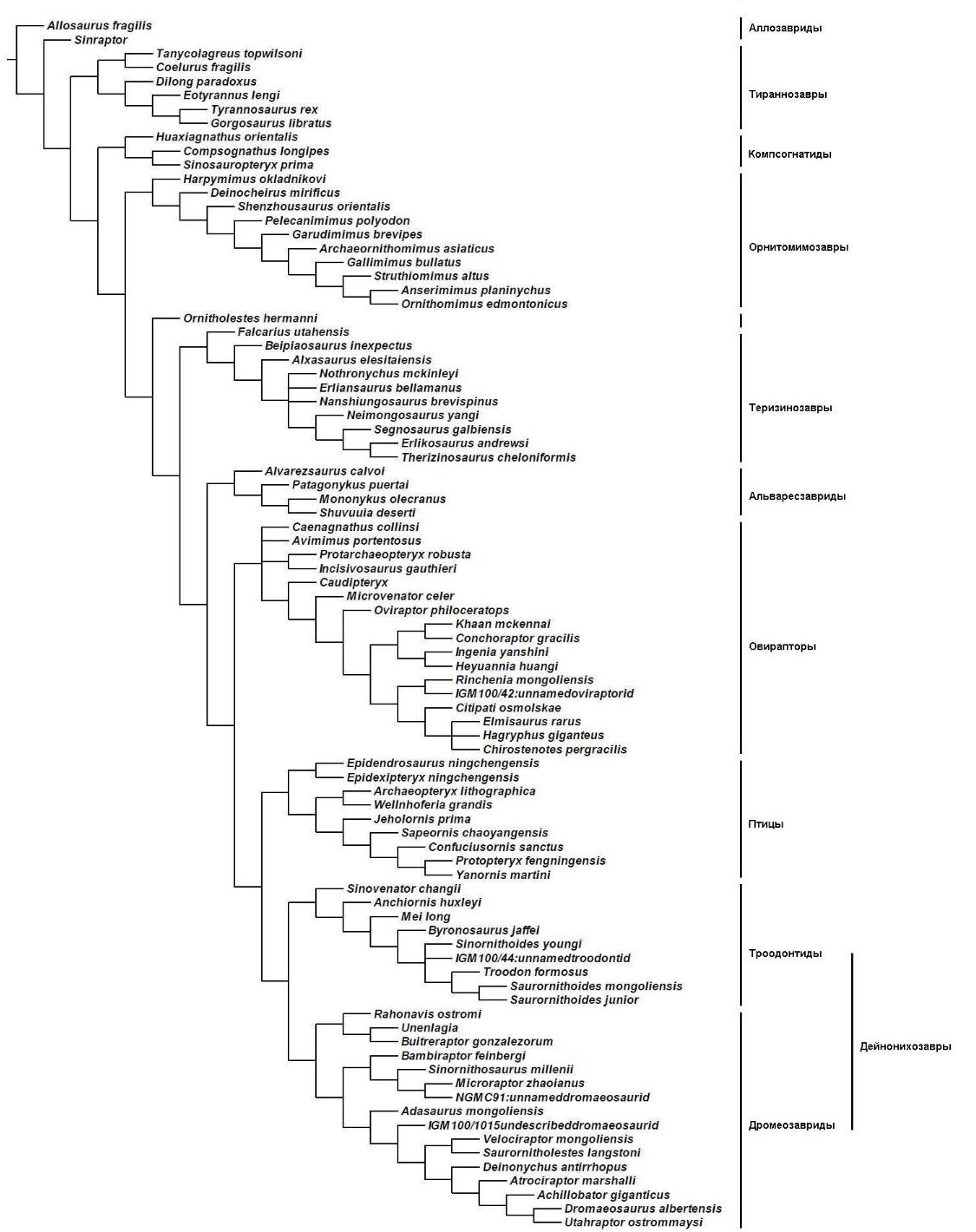
Итак, что мы можем предположить с уверенностью о еще не найденных переходных формах? Здесь пригодятся пояснения. В интересующей нас группе звероногих динозавров выделяют две подгруппы – карнозавров и целурозавров. Ранее к карнозаврам относили всех звероногих большого размера, а к целурозаврам – среднего и маленького размеров. Но с течением времени выяснилось, что огромные тираннозавриды анатомически принадлежат в большей мере к целурозаврам, чем к карнозаврам. Пришлось гипотезу об отличиях в размерах тела отбросить. Позднее выяснилось, что представители нескольких семейств целурозавров независимо увеличивались в размерах тела. Таким образом, целурозавры изначально маленькие животные. Это приводит к неизбежному заключению, что предок гигантского тираннозавра должен иметь скромный размер.
Строение передних конечностей предковой формы также можно без труда предвидеть. Тираннозавриды в анатомии передних конечностей сильно отклонились от своих родственников. Комбинируя информацию из разных семейств целурозавров, приходится заключить, что переходная форма к типичным тираннозавридам должна быть трехпалой, кости передних конечностей должны быть длинными и хорошо развитыми. Мы можем даже указать число фаланг пальцев. Это то же число, что у археоптерикса, дейнонихозавров и большинства других целурозавров – 2-3-4.
Арктометатарсус – еще одна особенность, которую мы не ожидаем встретить у предка тираннозаврид. Арктометатарсус известен в разных группах, но он не является, насколько мы знаем, наследственной особенностью целурозавров, так что мы не ожидаем его увидеть у ранних тираннозаврид.
Можно попытаться спрогнозировать и некоторые другие особенности, но это потребует более сложного анализа. Остановимся на достигнутом.
Так известны ли нам предки для семейства динозавров, которое захватило внимание даже далеких от палеонтологии людей? Тираннозавр был впервые описан в 1905 году, а его самый старый и хорошо сохранившийся на сегодняшний день предок – 101 год спустя. Останки динозавра, получившего имя гуанлонг, были раскопаны в нулевых годах XXI века на западе пустыни Гоби. Возраст отложений, в которых были найдены кости, составляет 160 миллионов лет, поздний юрский период. Анализ скелета со всей очевидностью показывает, что гуанлонг занимает промежуточное положение между ранними целурозаврами и тираннозавридами мелового периода. Он также подтверждает нашу эволюционную гипотезу о том, какими были предки тираннозаврид [90].
Гуанлонг в сравнении с более поздними тираннозаврами – небольшое животное, длинна тела ок. 3 метров. Кости передних конечностей не редуцированы, так что его передние конечности хорошо развитые и длинные (ок. 60% длинны задних). Три развитых пальца присутствуют, число фаланг соответствует типичному для целурозавров – 2-3-4. Второй палец не увеличен относительно других. Интересно, присутствует полулунная кость запястья. Ни у одного другого тираннозаврида во взрослом состоянии полулунная кость не обнаруживается в связи с редукцией передних конечностей.
Есть несколько других особенностей, которые не встречаются у тираннозаврид. Например, нижнечелюстное отверстие гуанлонга было большим в размере, у него также присутствовало тазовое отверстие.
Чтобы не усложнять описание, я сведу все особенности в таблицу.

Три особенности не попали в таблицу, так как их нельзя обозначить как "есть или нет". Они сами находятся в промежуточном состоянии. Это 1) Относительный размер межчелюстных зубов, 2) выступ на чешуйчатой и квадратно-скуловой костях, 3) положение подвздошных костей. У гуанлонга подвздошные кости сближаются относительно средней линии тела, но не входят в контакт, как у других тираннозаврид.
Гуанлонг не одинок в своем роде. Целурозавры подразделяются на несколько семейств, эволюционные отношения между которыми было очень трудно проследить из-за недостатка таких гуанлонгов. Но с течением времени искомые формы появились. Они подтвердили одни эволюционные гипотезы и опровергли другие. В 2009 китайские и американские палеонтологи подробно описали аналогичную гуанлонгу форму, на сей раз из семейства тех самых овирапторидов, охранявших свои гнезда. Динозавр получил имя инцизивозавр. Один абзац этого описания стоит процитировать полностью:
"Не считая известных пернатых динозавров, в джехольских отложениях были найдены бесчисленные переходные формы. Эти динозавры (например, лиаоцератопс, основные дейнонихозавры микрораптор и синовенатор, основной овирапторозавр каудиптерикс, переходной тираннозаврид дилонг, переходной теризинозаврид бэйпяозавр, основной орнитомимозавр шенцоузавр и ранний гадрозаврид Jinzhousaurus) обладают множеством особенностей, которые помещают их в основе признанных эволюционных групп, и поэтому они имеют важнейшее значение для разработки точных филогенетических деревьев. Сказанное также верно для инцизивозавра, который последовательно размещается в основе древа овирапторазавров".
Balanoff A. et al. "Cranial osteology of the theropod dinosaur Incisivosaurus gauthieri (Theropoda: Oviraptorosauria)" [3]. Внутренние ссылки опущены.
Заинтересованным читателям я предлагаю самостоятельно изучить анатомию упомянутых динозавров, а также признаки их семейств, создать сравнительные таблицы и установить самим, являются ли перечисленные животные переходными формами или нет. Впрочем, эта работа уже проделана на профессиональном уровне. Палеонтологи, используя теорию вероятностей, вычислили эволюционные отношения между целурозаврами, определяя степень неслучайного совпадения анатомических черт.

Но, может быть, я просто пытаюсь выдать желаемое за действительное? Правда ли, что все переходные формы среди целурозавров (и вообще среди позвоночных животных) уже найдены, и палеонтологам осталось лишь вставлять новые роды в установленную схему? Нет. Хотя некоторые эволюционные отношения среди целурозавров, как думают, наконец-то надежно установлены, пробелы в нашем знании не исчезают все сразу. Связи между поздними (меловой период) целурозаврами стали более или менее ясными. Но ранняя эволюция целурозавров плохо изучена. Как сегодня считается (и это еще одна гипотеза), корни известных семейств следует искать в отложениях не позднее середины юрского периода. Примечательно, что до настоящего времени палеонтологам известно сравнительно немного хорошо сохранившихся динозавров этого временного интервала. Если где-то существует формация, хранящая изобилие экземпляров динозавров середины Юрского периода, то пока ее никто не обнаружил. Остается лишь надеяться на подобное открытие. Но палеонтология – "медленная" наука.
Также можно указать трудности другого рода. Из приведенной выше кладограммы следует, что семейство альваресзаврид ближе связано с семейством теризинозаврид. Альваресзавриды – очень специализированные динозавры, своей анатомией бросившие вызов фантазии палеонтологов. Кто может быть их родственником – палеонтологическая загадка, в сравнении с которой меркнут даже дзенские коаны. С кем только альваресзаврид не роднили. В них видели даже птиц, утративших способность к полету – мезозойских страусов. Пока что переходная форма от примитивных целурозавров к альваресзавридам не найдена. Когда она будет найдена (а никто из палеонтологов не сомневается в такой возможности), можно будет проверить, правильно ли палеонтологи оценили совпадения анатомических черт альваресзаврид и теризинозаврид, действительно ли они – двоюродные братья.
Более того, представленная выше схема эволюционных отношений также является гипотезой. Она неизбежно должна подвергаться проверке по мере описания новых окаменелостей и переоценки старых.
UPD: Искомый альваресзаврид был описан во время подготовки этой статьи. Я не стал переписывать первоначальный текст для иллюстрации того, как могут устаревать данные. Динозавр гаплочериус частично подтвердил связь альваресзаврид с теризинозавридами. Поскольку гаплочериус один из самых ранних целурозавров, я также добавил его в таблицу для сравнения с близкими родственниками птиц [14].








{kind=link}
{kind=link}
{kind=link}
Оценили 19 человек
34 кармы