

Да , да ! Именно так дамы и господа ! Вот уже много времени мы можем быть очевидцами знаменательного события в истории жизни на земле - " годовщины победы на рептилиями " .
Это факт , коей на все сто процентов достоверен и не подлежит сомнению .
Но что бы понять , что же случилось начнем пожалуй с самого начала .
И так , жизнь на земле зародилась 3,6 миллиардов лет назад , но первые многоклеточные появились только около миллиарда лет назад . Это были существа вроде медуз и морских ежей .
Пример : Чарния (лат. Charnia, от названия возвышенности Чарнвуд-Форест[en]) — род организмов эдиакарского периода неясной классификации. Известны по отпечаткам продолговатой формы с зигзагообразно расположенными сегментами. Род представлен 3 видами.
Обнаружение отпечатков чарнии было важным шагом в палеонтологии, поскольку до 1970-х годов предполагалось, что в протерозое вообще не было крупных живых организмов, а лишь микроорганизмы (обнаруженные в 1930-х годах в Намибии и 1940-х годах в Австралии подобные отпечатки ошибочно относили к кембрию).

Но для нас в первую очередь важны сущейства вроде пикайи .
Пикайя (лат. Pikaia gracilens) — небольшое примитивное хордовое животное. Ископаемые останки пикайи встречаются в среднекембрийских отложениях возрастом 530 млн лет в сланцах Бёрджес (Канада).

Длина тела взрослых особей достигала от 1,5 до 6 см, в среднем она равнялась 4 см. Высота тела составляла от 7 до 16 % его длины, в среднем около 11 %.
Тело пикайи удлинённой формы, округлого сечения в передней части и сжатое с боков в остальных частях. Вдоль брюшной стороны тянулся узкий гребень, напоминающий плавник. Еще один гребень, хотя и не столь высокий, располагался на спинной стороне.
Голова была очень маленькая, подразделенная на две округлые лопасти, на которых имелась пара щупалец. Есть основания полагать, что эти щупальца были упругими, поскольку после смерти животного они обычно оставались прямыми, судя по сохранившимся отпечаткам. Скорее всего, они выполняли функцию органов обоняния и осязания. У основания головы на её нижней стороне размещалось ротовое отверстие. Никаких органов, похожих на глаза, не обнаружено.
Позади головы по обеим сторонам глотки имелось 9 пар ветвистых придатков, которые трактуются исследователями как наружные жабры. Вблизи оснований этих придатков иногда видны небольшие округлые пятна, возможно, соответствующие жаберным щелям.
Мускулатура ее разделена на множество поперечных сегментов — миомеров, которых насчитывается до сотни. Миомеры имеют сигмоидные очертания. Границы между миомерами хорошо заметны на ископаемых останках, поскольку в соответствующих местах отпечатков повышено содержание углерода. По всей видимости, миомеры были разделены прослойками плотной соединительной ткани, которая разлагалась медленнее, чем мускулатура животного. Сегментация распространяется и на брюшной плавник.
Внутри туловища на спинной стороне расположен плотный продольный тяж, который сохраняет объёмную, выпуклую форму даже у ископаемых останков. Он начинается позади глотки и тянется до самого конца туловища. Раньше он рассматривался исследователями как хорда. Однако, его толщина слишком велика для хорды. Кроме того, он расположен слишком высоко, не оставляя места для нервной трубки, которая должна размещаться выше, чем хорда. Прежняя интерпретация была пересмотрена в 2012 году С. Конвеем Моррисом и Ж.-Б. Кароном. В их работе этот тяж именуется спинным органом. Происхождение и функции данного органа пока остаются неясными .
Хорда и нервная трубка, по интерпретации С. Конвея Морриса и Ж.-Б. Карона, располагаются ниже спинного органа.
Пищеварительная система открывается ротовым отверстием у основания головы. В передней части кишечника имеется расширение — глотка. Анальное отверстие, согласно реконструкции С. Конвея Морриса и Ж.-Б. Карона, расположено на заднем конце тела.
Еще один тонкий продольный тяж расположен на брюшной стороне тела. С. Конвеем Моррисом и Ж.-Б. Кароном он интерпретируется как кровеносный сосуд. На некоторых экземплярах подобный сосуд обнаружен и на спинной стороне
В силурийский период из таких существ в значительной степени благодаря
ордовико-силурийскому массовому вымиранию из первых бесчелюстных развились первые челюстноротые - акантоды .
Аканто́ды, или колючкозу́бые(лат. Acanthodii, раньше — Acanthodei) — класс вымерших рыб. Согласно результатам последних исследований, акантоды представляют собой парафилетическую группу, относящуюся к стем-группе класса хрящевых рыб.
Название таксона восходит к имени входящего в него рода Acanthodes, первым получившего научное описание. Ранг данного таксона менялся; с 1940 года рассматривается как класс. Изучение акантодов ведёт своё начало с 1829 года, а в 1833—1845 годах в трудах Л. Агассиса было сформировано представление о них как о хорошо очерченной группе палеозойских рыб. С тех пор акантодов изучали многие палеонтологи, кардинально расширившие круг сведений об объёме таксона, строении и классификации акантодов. Ископаемые остатки акантодов обнаружены в отложениях начиная с середины силура и заканчивая концом перми; исчезновение акантодов связывают с массовым пермским вымиранием.
В своём строении акантоды сочетают признаки, характерные и для других групп палеозойских челюстноротых, отличаясь наличием хорошо выраженных плавниковых шипов перед всеми плавниками, кроме хвостового. Обитали как в морях, так и в пресных водоёмах; среди них преобладали планктонофаги и хищники. В составе класса выделяют четыре отряда, а также ряд родов неясного систематического положения .

Ордовикско-силурийское вымирание (англ. Ordovician extinction — «ордовикское вымирание») — массовое вымирание на границе ордовикского и силурийского периода, около 450—440 миллионов лет назад. Третье по процентной части вымерших родов из пяти сильнейших вымираний в истории Земли и второе — по потерям в количестве живых организмов.
В период между 450 и 440 млн лет назад, с промежутком в 1 млн лет, произошли два всплеска вымирания. Для морских организмов это вымирание второе по величине, оно уступает только пермскому вымиранию. В то время вся известная жизнь обитала в морях и океанах. Погибло более 60% морских беспозвоночных, включая две трети всех семейств брахиопод и мшанок. Особенно пострадали брахиоподы, двустворчатые моллюски, иглокожие, мшанки и кораллы. Непосредственной причиной вымирания, как представляется, явилось движение Гондваны к области южного полюса. Это привело к глобальному похолоданию, оледенению и последовавшему за ним падению уровня мирового океана. Отступление границы океана разрушило или повредило места обитания вдоль континентального побережья. Данные об оледенении были найдены в отложениях в пустыне Сахара. Комбинация низкого уровня мирового океана, похолодания и образования ледников, вероятно, и послужила причиной ордовикского вымирания. Эта была лишь часть одного более глобального события под название гуронское оледенение и земля - снежок .
«Земля-снежок» (англ. Snowball Earth) — гипотеза, предполагающая, что Земля была полностью покрыта льдом в части криогенийского и эдиакарского периодов неопротерозойской эры, а также, возможно, в другие геологические эпохи. Гипотеза призвана объяснить отложения ледниковых осадков в тропических широтах во время криогения (850—630 млн лет назад) и другие загадочные черты геологической летописи криогения. После окончания последнего большого оледенения ускорилась эволюция многоклеточных.Причиной этого события были химические реакции в атмосфере .Соприкасаясь с воздухом, силикатные горные породы подвергаются химическому выветриванию, которое удаляет диоксид углерода из атмосферы. В общем виде эти реакции выглядят так: минерал + CO2 + H2O → катионы + бикарбонат + SiO2. Пример подобной реакции — выветривание волластонита:
CaSiO3 +2 CO2 +2 H2O → Ca2+ + SiO2 + 2 HCO3−
Освобождённые катионы кальция реагируют с растворённым бикарбонатом в океане, образуя карбонат кальция как химически осаждённую горную породу. Это переводит углекислый газ из воздуха в литосферу, и в стабильном состоянии на геологических масштабах времени компенсирует выделение углекислого газа вулканами.
Когда Земля охлаждается (из-за естественных климатических флуктуаций и изменений в солнечной радиации), скорость химических реакций падает, и этот тип выветривания замедляется. В результате меньшее количество диоксида углерода извлекается из атмосферы. Повышение концентрации диоксида углерода, являющегося парниковым газом, приводит к обратному эффекту — Земля разогревается. Эта отрицательная обратная связь лимитирует силу похолодания. Во времена криогения все континенты были в тропиках возле экватора, что делало этот сдерживающий процесс менее эффективным, так как высокая скорость выветривания сохранялась на суше даже во время охлаждения Земли. Это позволило ледникам продвинуться далеко от полярных регионов. Когда ледник продвинулся достаточно близко к экватору, положительная обратная связь через увеличение отражательной способности (альбедо) привела к дальнейшему похолоданию, пока Земля целиком не обледенела. Не менее грандиозными были более ранние оледенения, такие как Гуронское оледенение.
Гуро́нское оледене́ние — одно из древнейших и наиболее продолжительных оледенений на Земле. Началось и закончилось в палеопротерозое и длилось около 300 млн лет. Причиной гуронского оледенения была кислородная катастрофа, в ходе которой в атмосферу Земли поступило большое количество кислорода, выработанного фотосинтезирующими организмами. Метан, который ранее присутствовал в атмосфере в больших количествах и давал основной вклад в парниковый эффект, соединился с кислородом и превратился в углекислый газ и воду ( метан в 28 раз более сильный ,парниковый газ чем углекислый газ ) Изменения состава атмосферы, в свою очередь, привели к сокращению численности метаногенов, что вызвало дополнительное снижение уровня метана.

Колоссальные масштаб и длительность гуронского оледенения могут быть связаны и с так называемым парадоксом слабого молодого Солнца.

Парадокс слабого молодого Солнца — наблюдаемое противоречие между палеоклиматическими данными и астрофизическими моделями эволюции Солнца.
Стандартная модель эволюции звёзд утверждает, что 4 млрд лет назад Солнце излучало приблизительно на 30 % меньше энергии, чем сейчас. При таких условиях вода на поверхности Земли должна была бы полностью замёрзнуть. В то же время геологические исследования архейских осадочных пород показывают, что в эту эпоху на Земле был влажный и тёплый климат. В условиях глобального оледенения, возможно, не смогла бы возникнуть жизнь.
Большинство учёных склоняются к объяснению этого парадокса глобальным парниковым эффектом, действовавшим в ранней истории Земли и вызванным очень высокими концентрациями таких вулканических газов, как углекислый газ и метан. Впервые эту модель предложили и количественно анализировали советские ученые Л. М. Мухин и В. И. Мороз. В роли парникового газа могла выступать смесь азота и водорода, которого в атмосфере молодой Земли было больше, чем сейчас. Альтернативные гипотезы, объясняющие достаточный для зарождения и поддержания жизни приток тепла, включают изменение структуры Солнечной системы, при котором Земля родилась ближе к Солнцу, и изменения в расположении и площади материков планеты.
В работах разных палеогляциологов хронологические рамки указаны по-разному. Согласно одной из версий, оледенение началось в сидерии 2,4 млрд лет назад и закончилось в конце риасия, 2,1 млрд лет назад. В работах же ряда других гляциологов (в частности, Дугласа Бенна и Дэвида Эванса) период фигурирует как Макганьенское оледенение, в честь формации Макганьен в Южной Африке.
Но восновном все это было вызвано чисто биотическими причинами - замещением старых групп новыми . К примеру доминирующая до него группа головоногих была замещена членистоногими .
Так вот , рыбы изначально были редки и незначительны , но к девонскому периоду они стали наиболее развитой формой жизни . Доминирования достигли панцерные рыбы - плакодермы .
Плакоде́рмы или пластинокожие рыбы (устар. панцирные рыбы, лат. Placodermi, от др.-греч. πλᾰκός «плита» и δέρμα «кожа») — класс челюстноротых (Gnathostomata), живших в палеозое, начиная с раннего девона. Расцвет пластинокожих рыб пришёлся на этот же период, а в конце карбона группа полностью вымерла. Ранние плакодермы жили исключительно в морской воде, но позже переселились и в солоноватоводные водоёмы.
Термин «панцирные рыбы», часто применяемый для класса в старой литературе, является некорректным и не отражает особенностей анатомии плакодерм: их так называемый «панцирь» на самом деле представлял собой набор пластин, сверху покрытых кожей, а не роговыми щитками .

Характерным признаком плакодерм была защищённость головы и туловища кожными (дермальными) пластинками. Самых крупных размеров достигли представители отряда артродир: хищный дунклеостей, достигавший 10 метров в длину, планктонофаги Homostius и Titanichthys до 8—9 метров в длину.
Плакодермы были одними из первых позвоночных, у которых развились челюсти (предполагается, что эти челюсти развились из жаберных дуг бесчелюстных рыбообразных). При этом строение челюстей у плакодерм сильно отличается от строения челюстей у современных рыб. Так, у современных рыб эндохондральный нёбноквадратный элемент верхней челюсти расположен медиальнее основных мышц, смыкающих челюсти; у примитивных плакодерм он тесно связан со щекой, находясь латеральнее пространства, занимаемого челюстной мускулатурой. У большинства плакодерм вместо зубов к краям челюстей прикреплены крупные покровные костные пластины, причём ни у одного из видов неизвестно регулярной смены зубов, обычной для других рыб. Эти особенности позволяют предполагать, что плакодермы произошли от линии примитивных челюстноротых позвоночных, отличных от форм, которые дали начало современным хрящевым и костным рыбам.
Все роды плакодерм обладали грудными и брюшными плавниками, но ни у одного не известен анальный. Хвост преимущественно гетероцеркальный (верхняя его лопасть, в которую заходит позвоночник, больше нижней).
Наши предки к тому времени готовились к выходу на сушу .
Знакомтесь : лопастиперые рыбы .
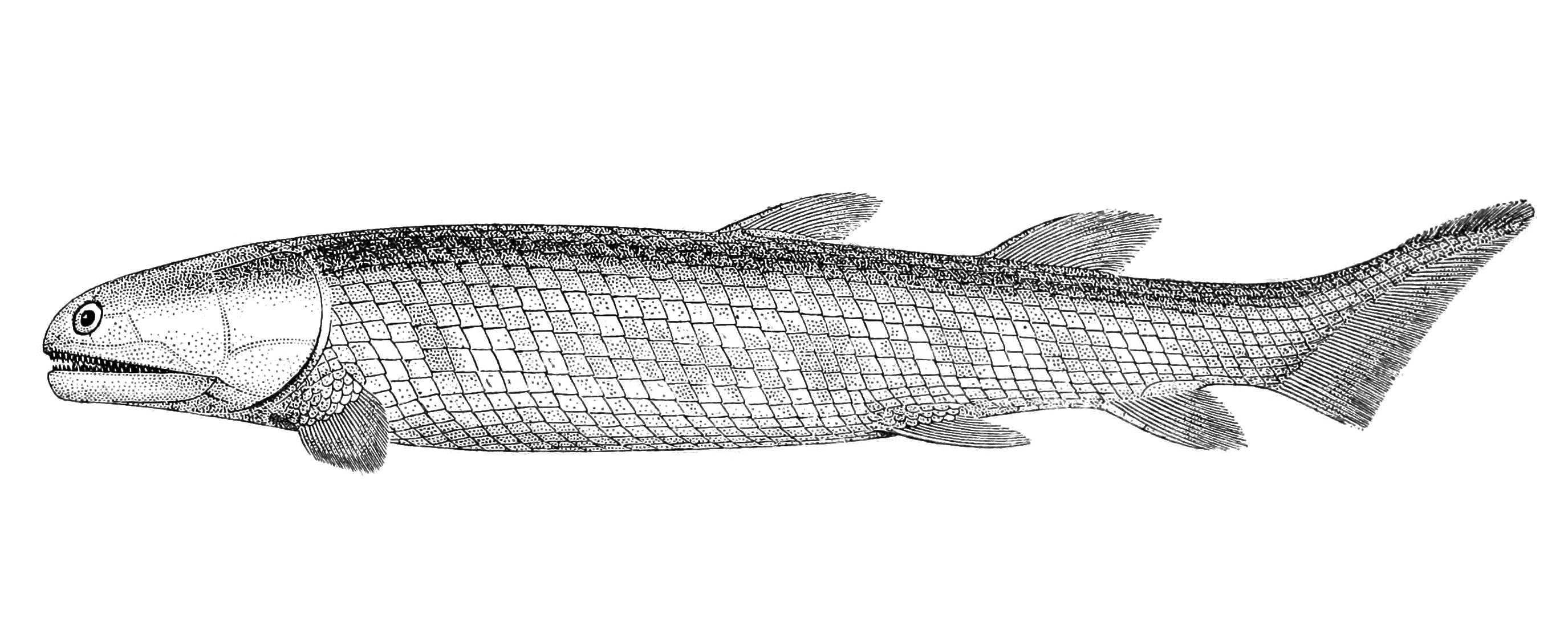
Лопастепёрые рыбы, или мясистолопастные, или хоа́новые(лат. Sarcopterygii) — класс костных рыб. Известны с раннего девона. Современные лопастепёрые относятся к отрядам целакантообразные и рогозубообразные.
Но особенно интересено существо под названием тиктаалик.

Тиктаа́лик (лат. Tiktaalik roseae) — вид ископаемых лопастепёрых рыб из верхнего девона (франский век), имевших много общих черт с четвероногими. Название означает «налим» на языке инуитов, населяющих арктическую Канаду. Ископаемые останки были обнаружены в 2004 году в отложениях позднего девона (около 380 млн лет назад) на острове Элсмир (территория Нунавут, север Канады) палеонтологами Эдвардом Дешлером, Нилом Шубиным и Фаришем Дженкинсом.
Тиктаалик является переходным звеном между рыбами и наземными позвоночными. В его строении сочетаются черты тех и других.
Признаки, свойственные рыбам: жабры и чешуя.
Признаки, характерные для переходных форм от рыб к четвероногим: строение костей и суставов конечностей, в том числе функциональный лучезапястный сустав, свойственный четвероногим, при наличии плавников, а также строение уха.
Признаки, традиционно приписываемые четвероногим: рёбра, по строению схожие с рёбрами четвероногих; подвижный шейный отдел; лёгкие.
Именно на этом уровне , либо на уровне амфибий происходит разделение на ветви амниот : парарептилий , тероморфов и завроморфов .
Парарептилии (лат. Parareptilia) — сборная, возможно, искусственная, группа примитивных четвероногих. Термин предложен Э. Олсоном в 1947 году. Различные классификации дают разный состав для этой группы. Так, согласно М. Ф. Ивахненко (2001), парарептилии — подкласс в классе рептилиоморф (Reptiliomorpha). В данной схеме в состав парарептилий входят сеймуриаморфы, проколофономорфы и хелониаморфы (надотряд Cheloniamorpha). В последний надотряд входили никтеролетеры, токозавры, лантанозухи, рипейазавры, парейазавры и черепахи. В целом в этой классификации парарептилии выглядят естественной группой, возникшей от амфибийных (анамний) предков-сеймуриаморф, независимо достигшей амниотного уровня и приведшей в итоге к появлению современных черепах.
В западных кладистических классификациях парарептилии принимаются как подкласс рептилий (то есть включают только амниот) и приравниваются к анапсидам. В таком варианте группа включает в себя диадектов, лимносцелисов, болозавров, мезозавров, миллеретид, все группы высших «парарептилий» по М. Ф. Ивахненко, а также, в ряде классификаций — капторинид. Сеймуриаморфы исключены из группы. Черепахи могут и не принадлежать к парарептилиям. С другой стороны, ряд групп анапсид (парарептилий) близки к основанию ствола завропсид. Поэтому результаты молекулярных исследований, сближающих черепах с диапсидами, не являются противоречивыми (кроме того, черепахи — единственные современные «анапсиды», логично, что они будут близки всё-таки к завропсидной ветви, а не к синапсидам).
Пример :никтифрурет
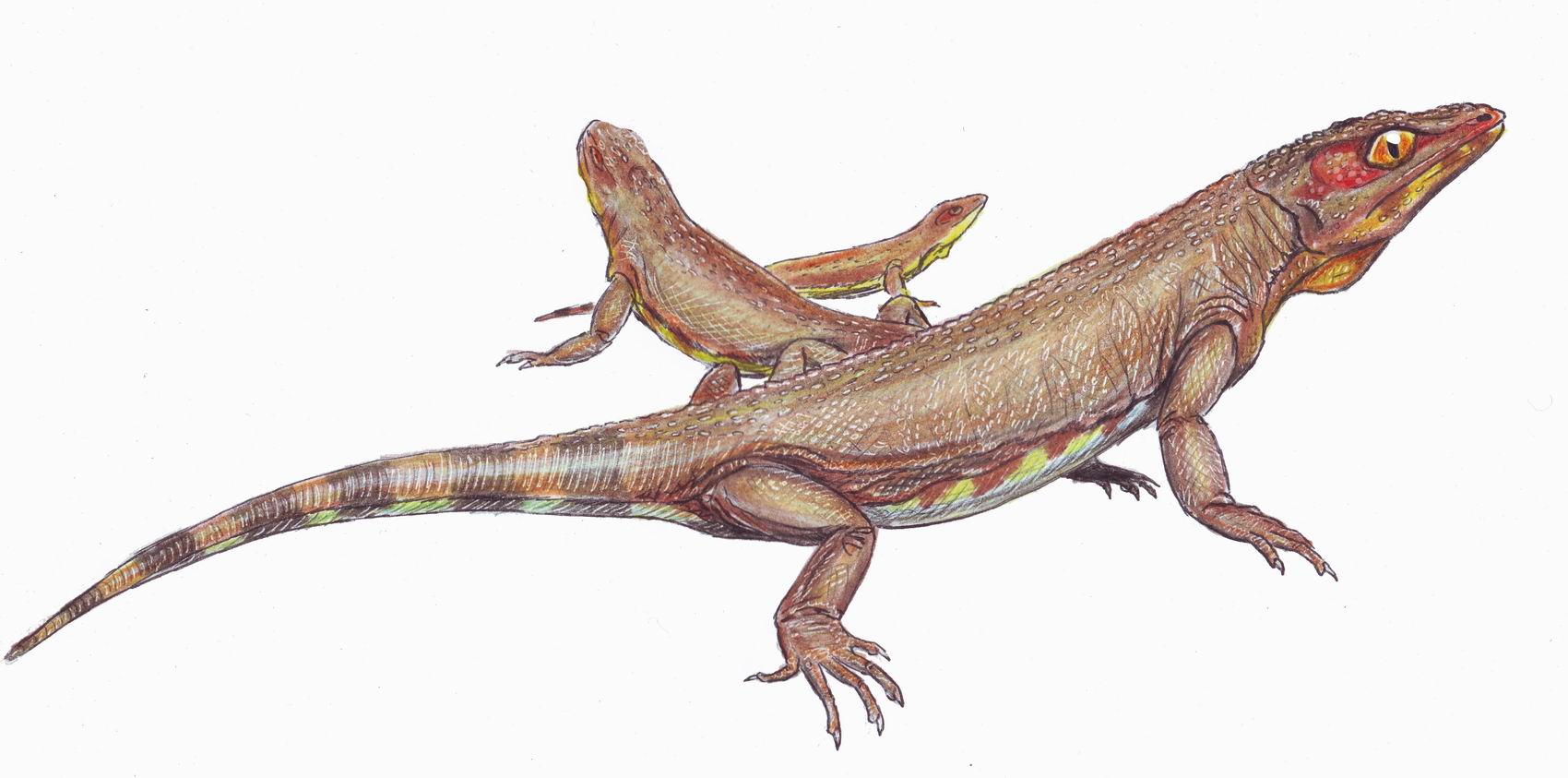
Никтифрурет (лат. Nyctiphruretus, от др.-греч. νυκτιφρούρητος «несущий ночную стражу») — род вымерших примитивных рептилий верхнепермской эпохи. Принадлежит к отряду проколофономорфов подкласса анапсид. Описан И. А. Ефремовым в 1938 году из «среднепермских» отложений реки Мезень.
Мелкое животное, длиной около 30—40 см, внешне похож на ящерицу. Глазницы огромные (вероятно, глазница расширена назад для крепления челюстной мускулатуры), большое отверстие теменного глаза. Возможно, имелась солевая железа возле ноздри. Ефремов считал никтифрурета ночным животным (отсюда название). Крыша черепа покрыта мелкими остеодермами — это предполагает наличие чешуи.
Не исключено, что никтифрурет был растительноядным и питался водорослями (строение его зубов сходно с таковым у никтеролетера). С другой стороны, он мог питаться насекомыми. Характерен для мезенского субкомплекса «средней» перми Приуралья, составляет до 75 % всей фауны. Обитал в каламитовых зарослях по берегам соленых водоемов, по образу жизни мог напоминать игуан-амблиринхов. Интересно, что молодью никтифруретов питалась крупная парарептилия макролетер (Macroleter). Типовой вид — Nyctiphruretus acudens. Второй вид — Nyctiphruretus optabilis — описан В. В. Булановым в 2002 году.
Следующая ветвь - тероморфы или синапсиды , наши прямые предки .
Синапси́ды (лат. Synapsida), также тероморфы или зверообразные (лат. Theromorpha, от др.-греч. θήρ, θηρίον ‘зверь’ и μορφή ‘форма’) — таксономическая группа в составе амниот, возникшая около 318 млн лет назад[4]. Одна из групп синапсид — цинодонты (Cynodontia) — была предком млекопитающих[5][6].
На протяжении большей части XX века синапсиды считались одним из подклассов класса пресмыкающихся. Так, в монографии Р. Кэрролла[7] синапсиды выступали именно как подкласс класса пресмыкающиеся (Reptilia), включавший 2 отряда: пеликозавры (Pelycosauria) и терапсиды (Therapsida).
Позднее широкое распространение представлений, характерных для филогенетической систематики, привело к пересмотру классификации синапсид. Они были выведены из состава класса пресмыкающихся, получив в конце XX века статус самостоятельного класса.
По современным представлениям, синапсиды вместе с млекопитающими образуют кладу Synapsidomorpha (синапсидообразные), которая вместе с сестринской группой Sauropsida (завропсиды) образует, в свою очередь, кладу амниот — крупнейшую по объёму группу в составе надкласса четвероногих. При этом сторонники эволюционной таксономии выделяют в составе синапсидообразных два класса: парафилетический класс Synapsida и дочерний монофилетический класс Mammalia. Сторонники же кладистики (для которой характерно стремление избавиться от немонофилетических таксонов) обычно не выделяют класс синапсид, трактуя входящие в него группы независимо, а термин «Synapsida» нередко используют взамен термина «Synapsidomorpha» (при таком подходе понятие «синапсиды» охватывает и утконоса, и человека).
Пример : диметродон

Диметродо́н (лат. Dimetrodon) — род вымерших хищных синапсид из семейства сфенакодонтов, живших в пермский период 285—265 млн. лет назад. Эволюционно находится ближе к млекопитающим, чем к истинным рептилиям. Это крупнейший хищник своего времени .Достигал в длину 2,7—4,0 метров и был крупнейшим хищником своего времени. В отличие от рептилий имел зубы двух типов (резцы и клыки), однако не подобранные под туловище ноги. Наиболее яркой чертой диметродона является парус из натянутой на выросты спинных позвонков кожи. Подобные паруса развились у различных доисторических животных (амфибии Platyhystrix, пеликозавра Edaphosaurus, динозавра Spinosaurus) и служили терморегуляторами. Подсчитано, что 200-килограммовый холоднокровный диметродон разогревался бы без паруса с 26 °C до 32 °C за 205 минут, а с парусом — за 80 минут. По другим версиям парус использовался в брачных играх, служил камуфляжем среди вертикальных стеблей растений или настоящим парусом при плавании. «Парус» развивался постепенно в течение жизни, известны находки молодых особей диметродона с невысокими остистыми отростками. Молодые особи, по-видимому, обитали в зарослях по берегам водоёмов, взрослые — жили в лесах и на равнинах. Тем не менее, бытующее в популярных книгах изображение диметродона на фоне пустыни представляется нереальным — не будучи настоящей рептилией, он не мог бы выжить в жаркой сухой местности.
Диметродоны питались любыми животными, которых могли одолеть. Интересно, что в условиях примерно равного соотношения биомассы хищников и растительноядных (характерного для раннепермских фаун), основу питания в отдельные периоды могла составлять рыба. В 2008 году команда Р. Бэккера обнаружила четкие признаки того, что диметродоны поедали пресноводных акул — ксенакантид. Возможно, акулы оставались в пересыхающих прудах, где становились жертвой пеликозавров .
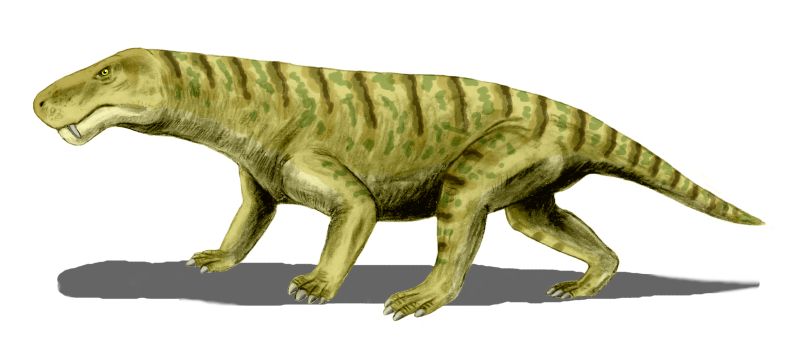
Первыми настоящими истинными сверхищниками суши был тоже наш поцан , терапсид , тоесть потомок пеликозавров вроде диметродона . Это был могучий и огромный по нашим меркам хищник -
Иностра́нцевия (лат. Inostrancevia) — представитель хищных терапсид из группы териодонты — горгонопсов. Вероятно, самый крупный из хорошо известных горгонопсов. Тело вытянутое, сжатое с боков. Мощный хвост. Череп узкий и удлинённый. Височные окна крупные, вытянуты вверх-назад. Орбиты средних размеров, вынесены наверх. Нёбные бугры и нёбные зубы отсутствуют. Позвоночник лёгкий, подвижный. Иностранецевия является первым из гигантским хищников в истории Земли на суше . По размерам она была сопоставима с саблезубыми кошками.
Впервые открыт В. П. Амалицким в 1898 году. Найдены иностранцевии в верхнепермских отложениях на берегах реки Северной Двины и в Оренбургской области. Название дано в честь русского геолога А. А. Иностранцева. Вместе с православлевией (Pravoslavlevia) выделяется в подсемейство Inostranceviidae (англ.)русск., близкое к рубиджеидам (Rubidgeinae (англ.)русск.).
Близ Очёра поставлен памятник иностранцевии. Увидите памятник - отдайте ему честь . Это наш братан и не простой , а прославиший род тероморфов .
Вот так это животное выглядело :
Но случилось страшное - в вязниковском ярусе(Геологи́ческий я́рус — стратиграфическое подразделение, единица общей стратиграфической шкалы, подчинённая геологическому отделу. Подразделяется на стратиграфические зоны. Объединяет толщу горных пород, образовавшуюся в течение одного геологического века (как правило, несколько миллионов лет) и отвечающего определённому этапу геологического развития Земли. Характеризуется типичными для него и только ему свойственными родами, подродами и группами видов. Решающим критерием для установления объёма и границ яруса являются данные биостратиграфического анализа.) ( если проще , то если брать период как век , то ярус-это год ) пермского периода существа подобные иностроцевии , тигры своего времени ( горгонопсы ) вымерли .
И на их место вылезло животное под названием архозавр .
Архозавр (Archosaurus rossicus) — единственный позднепермский представитель группы протерозухий , предков эритозухий и соответственно и куроротазов с авимитарсаллиями . . Описан Л. П. Татариновым из самых верхних горизонтов перми Владимирской области (Вязники) в 1960 году. Череп относительно высокий, зубы мощные, слабо изогнутые, длина черепа до 40 см, общая длина взрослых особей до 2 метров. Это животное было наземным охотником за крупной добычей. В вязниковском комплексе нет горгонопсов и архозавр занял место крупнейшего хищника — охотника за парейазаврами и дицинодонтами. Неописанные протерозухии указываются и для поздней перми ЮАР.
Вот так эта животина выглядела :
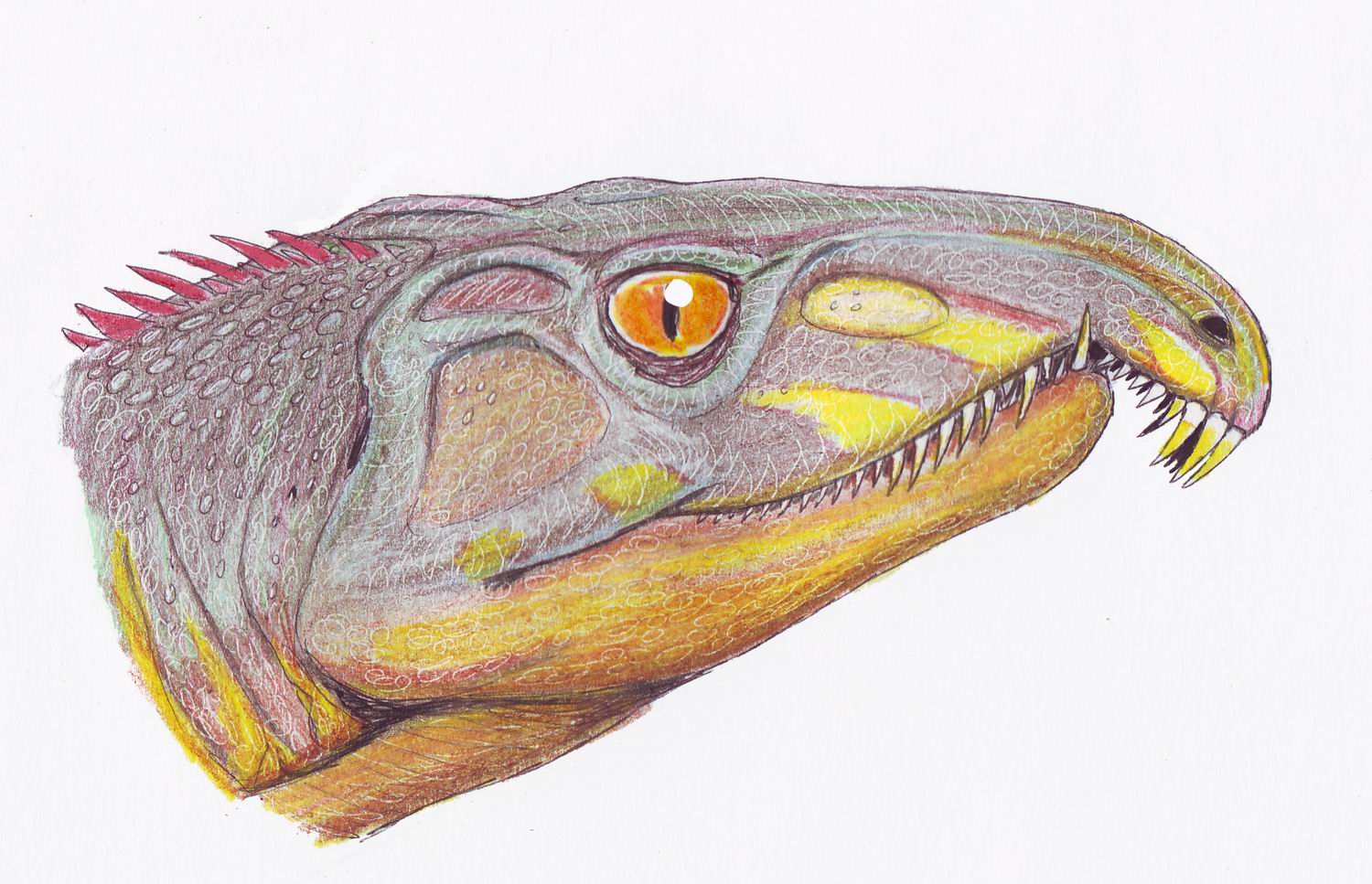
Она как я уже говорил принадлежит к семейству протерозухий . Эти существа занимали нишу крокодилов и варанов своего времени .
Протерозухии (лат. Proterosuchidae) — семейство пресмыкающихся из группы Archosauriformes, находятся в основании ветви, ведущей к более развитым истинным архозаврам. По устаревшей классификации являлись подотрядом «текодонтов» Proterosuchia Broom, 1906.
Впервые появились в конце пермского периода, вымерли в триасовом периоде (252,3—242,0 млн лет назад) .
А вот кто такие круротарзы и авиамитарсаллии с эритозухиями и что было дальше вы узнаете в следующей моей статье , а заодно и про всех завроморфов .




Оценили 2 человека
2 кармы