

Иногда, когда солнышко скроется за тучкой и изжога одолеет, лезут в голову грустные мысли, что миновали, дескать, времена Индиана Джонсов, когда можно было зайти в пещеру и узреть целый скелет неведомого существа, картинно раскинувшийся на полу. Ан нет, не прошли! Разве что теперь для этого надо быть тощим.
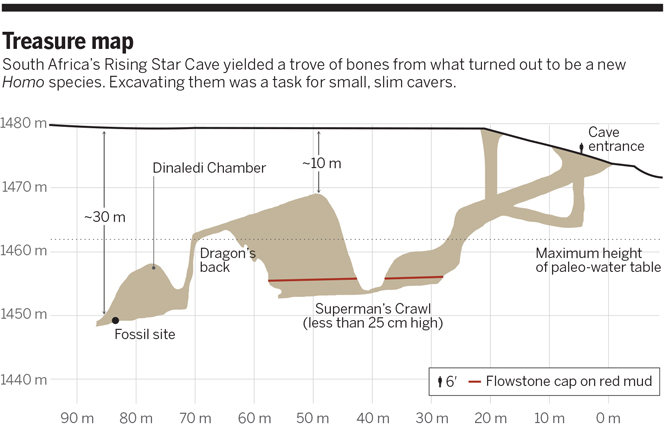
13 сентября 2013 г. два спортсмена-спелеолога – Стивен Такер и Рик Хантер - залезли в пещеру Райзинг Стар в Южной Африке. Она уже была известна узким "шкуродёром" "путь Супермена", по которому могут проползти лишь исключительно субтильные личности. Но Стивену и Рику повезло: в конце изведанного ранее туннеля они обнаружили продолжение, ведущее в манящие недра земли. И они не зря последовали диггерскому зову - в тупике (который теперь зовётся Диналеди) их ждала сенсация. На дне небольшой камеры эффектно лежали чьи-то кости. Стивен и Рик оказались не просто спелеологами, а грамотными и образованными людьми. Они сфотографировали неожиданную находку и принесли фотографии Ли Бергеру, известному во всём мире южноафриканскому палеоантропологу. Несколько лет назад он прославился, открыв в пещере Малапа скелеты Australopithecus sediba. Очевидно, звезда удачи продолжает светить над его головой.
Ли Бергер организовал масштабную экспедицию в Райзинг Стар. Тут-то и обнаружилось, что иногда всё же полезно есть мало каши: очутиться на месте сенсации дано не каждому. Только самые мелкие исследователи способны протиснуться в каменные тиски и достичь вожделенных костей.
Но это всё присказка. Главное, конечно, не история открытия, а сами окаменелости.

И тут сенсация окончательно стала сенсацией.
Во-первых, находок оказалось не просто много, а невероятно много: в камере метр на девять обнаружилось более полутора тысяч фрагментов! По итогу тут нашёлся почти целый скелет и останки ещё четырнадцати особей - взрослых мужчин, женщин, подростков и детей. Часть костей покоилась в собранном положении. Например, на поверхности красовалась совершенно целая кисть, прямо как в малобюджетных фильмах ужасов. Такого ещё не бывало. Во всей Африке за век исследований найдено сопоставимое число окаменелостей. Более того, раскопки пока затронули лишь небольшой участок пола. Сколько ещё останков хранит пещера? Трудно себе представить...
Во-вторых, кости лежали просто на полу. Они были лишь слегка припорошены каменной шугой, которая тысячи лет оседала на них со стен и потолка. Всего насыпалось пятнадцать сантиметров осадка. Судя по всему, кости не были никем потревожены, разве что некими неведомыми заблудшими спелеологами, которые были не столь сознательны и никому не сообщили о находке, а только пошебуршили окаменелости. Геологические изыскания привели исследователей к мысли, что пещера всегда была тёмным закутком, в неё всегда вёл лишь тот же "шкуродёр" - тесный туннель, стены которого к тому же местами покрыты острыми выступами. В пещере нет никаких следов человеческой деятельности и ни одной кости крупных животных. Найдены лишь шесть птичьих костей и несколько грызунячьих резцов. Нет и отпечатков зубов хищников на останках, нет и следов орудий. Нет и следов переноса водой. Как же так? Ничего тут нет, кроме огромной кучи человеческих костей, попавших в пещеру в собранном виде. Как? Откуда? Осторожные исследователи полагают, что раньше в камеру вёл какой-то другой ход, через который люди (а также птицы и мыши) проникали сюда. Но как? Провалились случайно, как в Малапе? И так пятнадцать раз? И при этом ни один зверь крупнее мыши не повторил их печальной судьбы? К тому же, состояние осадков говорит за то, что накопление костей происходило некоторое время, оно не было одномоментным. Так что вряд ли сразу целая группа древних людей попала в западню.
Но геологи утверждают, что альтернативного пути вообще нет и не было.
И тут открывается великое поле для сенсационных предположений. Как и зачем люди заползали сюда? Конечно, они были мелкими, в этом смысле им было проще, чем нынешним спелеологам и антропологам, но не всё же определяется размерами. Они ползли на ощупь? Или - возможно ли такое?! - первобытные диггеры освещали себе путь факелами? Протискивались ли они в смертельный тупик, чтобы умереть тут спокойно? Или - а такое могло ли доселе придти кому-то в голову?! - затаскивали трупы почивших соплеменников? Чтобы оценить невероятность сих предположений - не просто смелых, а почти на грани фантастики - стоит ознакомиться с самими посетителями пещеры, обрётшими тут вечный покой.
И тут во весь рост встаёт проблема, прямо вытекающая из вышеуказанного отсутствия в Диналеди чего-либо, окромя человеческих костей: нет датировки. Обычно геологический возраст определяется, исходя из стратиграфии (последовательности осадков), состава фауны и флоры, радиометрическими способами - по вулканическим туфам, обожжённым костям или хотя бы песку. А тут нет ничего из этого. Даже песка!
Остаётся "в-третьих": морфология. Великое изобилие костей для любого антрополога - солнечный праздник с каруселями и мороженым.
Морфология не подвела. В принципе, в Диналеди могли обнаружиться останки хорошо известных австралопитеков или эректусов. Это было бы тоже здорово, но не так пафосно. А тут всё оказалось как надо - сенсационнее некуда. По итогу исследования группа антропологов описала новый вид древних людей - Homo naledi.
Чем же он уникален?
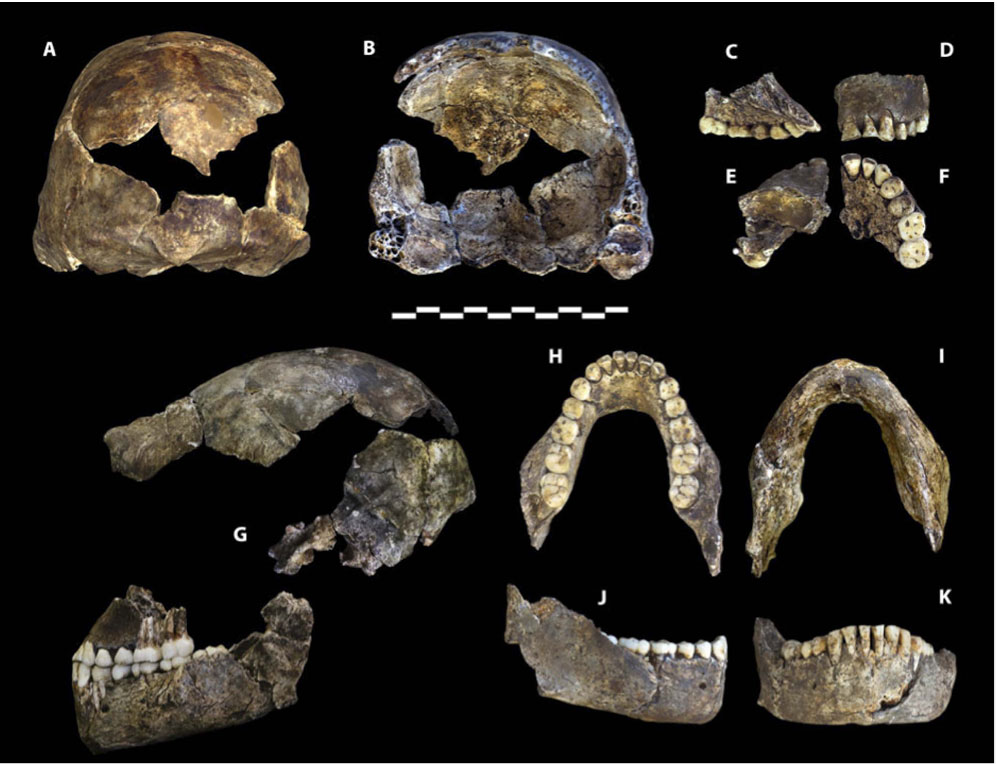
Диналедцы имели маленький рост - около полутора метров. Это не выделяет их из массы австралопитеков и "ранних Homo". Голова была тоже очень небольшой; мозгов было не то чтобы мало, но и не много - 465 см3 для двух маленьких черепов и 560 см3 для двух больших. Для австралопитеков бóльшая цифра - индивидуальный рекорд, для хабилисов меньшая - тоже, только теперь в минимальную сторону. По итогу, диналедцы занимают строго промежуточное положение. Это даже круче, чем в случае с седибой, чей объём - 420 см3, то есть скучно-австралопитековый.
Мозг Homo naledi выглядит весьма примитивно. Лобные доли низкие и короткие, хотя область "зоны Брока" - моторного речевого центра - вроде бы достаточно выпуклая. Особенно дремуче устроена височная доля: она фактически австралопитековая и особо не отличается от обезьяньего варианта. Лобная доля сходится с височной под тупым углом, как у шимпанзе.
Череп при этом не сказать, чтобы был чересчур примитивным. Мозговая коробка имеет наибольшую ширину в нижней части, а боковые стенки черепа при взгляде сзади несколько сходятся кверху, но не так уж резко. Относительная высота черепа вполне прилична по меркам "ранних Homo". Особенно интересно мощное развитие надбровных валиков и, соответственно, сильное заглазничное сужение. Такое строение больше подходит для классических эректусов, а не хабилисов. Впрочем, на женском черепе DH3 надбровные валики сравнительно тонкие. Лоб напрочь скошен, особенно на крупных черепах DH1 и DH2 - самцам и положено иметь брутальный вид. При этом на лобной кости прослеживаются лобные бугры: у обезьян и австралопитеков их не найти, а вот для Homo они вполне типичны. Височные линии выглядят даже прогрессивнее, чем у H. rudolfensis и H. habilis: они довольно слабые и следуют скорее по боковой, а не верхней стороне лобной кости. В задней части они не смыкаются с рельефом височной кости; зато имеется угловой торус - специфический элемент рельефа задней части теменной кости, типичный для эректусов и гейдельбергенсисов, но нехарактерный для "ранних Homo".
Затылок вполне австралопитековой конфигурации - укороченный и почти вертикальный, но с мощным наружным затылочным бугром, больше подходящим для эректусов.
На височной кости слуховые отверстия и сосцевидные отростки маленькие, примитивные. Впрочем, подавляющая часть тонкостей строения височной кости не отличает новый вид от ранее известных "ранних Homo".
О форме глазниц и носа остаётся больше догадываться. Возможно, новые материалы "откроют личико", но пока оно остаётся скромно завешенным тафономической вуалью. Впрочем, известно, что у наледи не было "передних лицевых валиков" - утолщений по бокам носового отверстия, типичных для южноафриканских австралопитеков.
Челюсти, очевидно, сильно выступали вперёд, особенно резко выражен альвеолярный прогнатизм на верхней челюсти. Передняя сторона нижней челюсти при этом не скошена, а почти вертикальна. Нёбо было мелким, чем Homo naledi больше похож на австралопитеков и H. habilis, нежели на более продвинутых людей, зато оно же было широким, - а это уже продвинутая черта. Альвеолярная дуга нижней челюсти самой что ни на есть умеренной формы: возможно, и не сапиентной, но точно и не австралопитековой. Удивительно прогрессивно выглядит высокое расположение подбородочного отверстия: таковое типично едва ли не только для современного человека.
Зубы Homo naledi - одна из самых продвинутых его частей.
Особенно удивительны маленькие размеры моляров. По ним диналедцы сопоставимы разве что с современным человеком. Они уступают не только всем австралопитекам и "ранним Homo", но вообще всем гоминидам, включая седиб и "хоббитов". Таких мелкозубых человечков с таким строением черепа никто найти не ожидал.
Столь же противоречиво и строение посткраниального скелета.
Позвоночник в большей степени напоминает вариант плейстоценовых Homo. Особенное внимание обращает на себя большой диаметр позвоночного канала. Грудная клетка широко расходится книзу - эта черта типична для австралопитеков. Столь же примитивны и лопатки. На плечевых костях почти отсутствует скрученность. Они очень грацильны, со слабо развитым мускульным рельефом, а локтевая с лучевой весьма прямы: вряд ли диналедцы лихо скакали по деревьям.
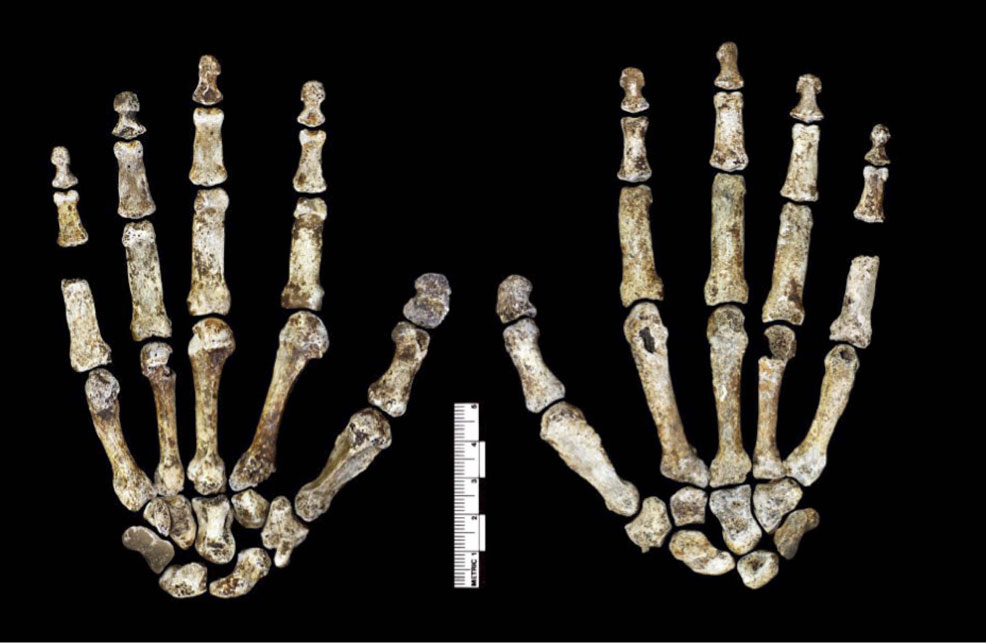
Кисть Homo naledi, с одной стороны, сохраняет удлинённость и примитивную изогнутость фаланг (даже бóльшую, чем у австралопитеков), с другой - имеет прогрессивное строение запястья и продвинутые пропорции пальцев. Например, большой палец действительно большой. Концевые фаланги расширены - это признак трудовой кисти. Одновременно, сустав кости-трапеции и первой пястной маленький, а на третьей пястной нет шиловидного отростка - это примитивные, отнюдь не трудовые черты. Есть и странности, скажем, форма первой пястной кости - с узким основанием и широкой головкой.
Тазовые кости сохранились не идеально, но ясно, что крылья подвздошной кости сильно расходились в стороны, как у австралопитеков. Вместе с тем, расстояние от седалищного бугра до вертлужной впадины уменьшено, как у A. sediba и Homo. Суммарное сочетание выглядит необычным.
Вдоль шейки бедренной кости тянутся какие-то странные гребни и бороздка - такого доселе не встречали ни у каких гоминид. Что это - локальная популяционная вариация или видовая специфика? Все образцовые признаки прямохождения - форма, длина и наклон шейки, выраженные ягодичная бугристость и шероховатая линия - развиты по полной программе.
На бугристости большой берцовой кости (Читатель может нащупать её у себя ниже коленной чашечки) имеется отдельный бугорок для сухожилия полусухожильной мышцы. Мышца эта сводит колени. Диналедцы часто держали пенальти?
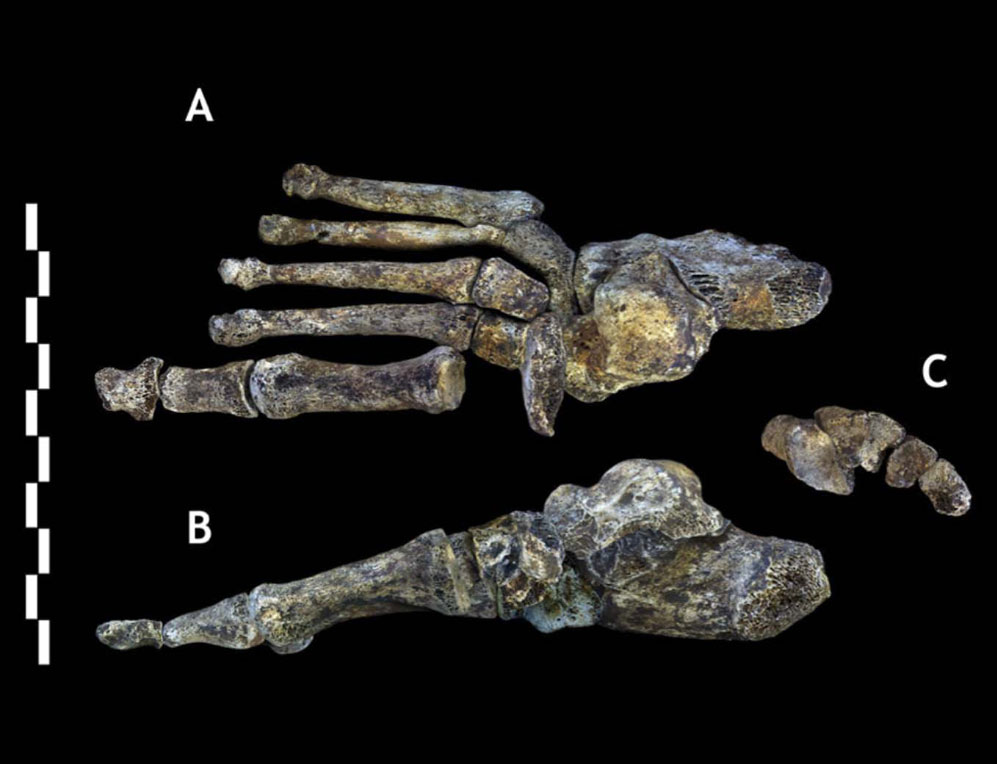
Стопа Homo naledi - едва ли не самая человечная его часть: на ней отлично развиты продольный и поперечный своды, нет никакого намёка на хватательные способности. При этом пяточный бугор выглядит уменьшенным, не вполне стандартны и некоторые частные пропорции вроде суженности основания первой плюсневой.
В целом, мы видим беспрецедентный набор странностей. Успокаивает хотя бы то, что разные индивиды минимально отличаются друг от друга, так что мы можем быть уверены, что все они принадлежат к одному виду, а их особенности действительно имеют видовой или популяционный характер, но не индивидуальный.
Как же в сумме объяснить всю эту мозаику?
В отсутствие датировки может быть несколько вариантов.
Во-первых, Homo naledi может быть действительно предком всех прочих людей. Комбинация примитивных и продвинутых черт запросто объясняется промежуточностью его эволюционного положения, а датировка может располагаться в интервале от трёх до двух миллионов лет назад. Однако, некоторые признаки наледи слишком прогрессивны. Скажем, трудно представить, чтобы крупные зубы австралопитеков превратились в столь маленькие Homo naledi, а после - опять в большие H. habilis и H. erectus (не говоря уж о мегадонтных H. rudolfensis), после чего опять стали уменьшаться. Не слишком ли много вывертов? В этом смысле линия австралопитеки - хабилисы - эректусы выглядит куда более последовательной, тогда как наледи и рудольфенсисы оказываются специализированными боковыми ответвлениями от генеральной линии.
Тут мы переходим к "во-вторых": Homo naledi может быть экзотической тупиковой ветвью эволюции, наряду с седибой, гари и парантропами. Одних уносило в увеличение зубов, других - в уменьшение. В этом случае Южная Африка представляется как место, где эволюция шла своим чередом, но этим людям не повезло: по совокупности признаков они проиграли гонку своим восточноафриканским родственникам.
Логично и "в-третьих": можно пофантазировать, что Homo naledi в действительности жили довольно поздно, одновременно с гораздо более продвинутыми гоминидами. Тогда они могли представлять реликтовый пережиток ранней стадии эволюции. В этом случае можно объяснить и мозаичное строение, и предполагаемое знакомство с огнём, и даже существование погребального обряда, хотя бы и крайне примитивного: дескать, подсмотрели у продвинутых кузенов, вот и собезьянничали, насколько позволяли австралопитековые мозги.
А может, правы скептики, и люди из Диналеди - это просто те же самые хабилисы, только мы наконец-то видим их собранный скелет?
Ведь до сих пор такой радости мы были лишены. Даже принадлежность кисти и стопы OH 7 и OH 8 - голотипов вида Homo habilis - к виду Homo habilis, как ни парадоксально, остаётся недоказанной. А вдруг OH 8 - это стопа бойсовского парантропа? Для подтверждения или опровержения нужны полные скелеты хабилиса и парантропа, а у нас нет ни того, ни другого. А в Диналеди мы можем иметь пусть своеобразно-мелкозубого и мелкоголового, но всё же просто хабилиса. Или это вариант того же седибы? Ведь он тоже сочетает признаки австралопитеков и хабилисов, пусть и в других комбинациях. Кстати, седиба также знаменит именно своей мозаичностью, он тоже весьма малоголов и мелкозуб, а по разным признакам таза, рук и ног может быть равно отнесён к австралопитекам или людям.
Всё же комплекс черт Homo naledi выглядит достаточно уникальным, чтобы признать его самостоятельным видом.
Ещё не все кости из Диналеди подняты на поверхность. Ещё не все их признаки описаны и осмыслены. Ещё многое будет о них сказано. Одно точно: наша генеалогия в очередной раз оказалась интереснее и богаче, а при рассказе о роде "Люди" отныне нельзя не вспоминать о "Человеке наледи".
Автор: Дробышевский С.
Источник: http://antropogenez.ru/
Материалы по теме:Источники:
Berger L.R., Hawks J., Ruiter de D.J., Churchill S.E., Schmid P., Delezene L.K., Kivell T.L., Garvin H.M., Williams S.A., DeSilva J.M., Skinner M.M., Musiba Ch.M., Cameron N., Holliday T.W., Harcourt-Smith W., Ackermann R.R., Bastir M., Bogin B., Bolter D., Brophy J., Cofran Z.D., Congdon K.A., Deane A.S., Dembo M., Drapeau M., Elliott M.C., Feuerriegel E.M., Garcia-Martinez D., Green D.J., Gurtov A., Irish J.D., Kruger A., Laird M.F., Marchi D., Meyer M.R., Nalla Sh., Negash E.W., Orr C.M., Radovcic D., Schroeder L., Scott J.E., Throckmorton Z., Tocheri M.W., VanSickle C., Walker Ch.S., Wei P. et Zipfel B. Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa // eLIFE, elifesciences.org, 2015, V.4, №e09560, pp.1-35.
Dirks P.H.G.M., Berger L.R., Roberts E.M., Kramers J.D., Hawks J., Randolph-Quinney P.S., Elliott M., Musiba Ch.M., Churchill S.E., Ruiter de D.J., Schmid P., Backwell L.R., Belyanin G.A., Boshoff P., Hunter K.L., Feuerriegel E.M., Gurtov A., Harrison J. du G., Hunter R., Kruger A., Morris H., Makhubela T.V., Peixotto B. et Tucker S. Geological and taphonomic contextf or the new hominin species Homo naledi from the Dinaledi Chamber, South Africa // eLIFE, elifesciences.org, 2015, V.4, №e09561, pp.1-37.
Тимирязев о Дарвине и происхождении нравственности.
Видеотека. Эволюция от обезьяны к человеку.
О природе человека: происхождение человека в свете новых антропологических находок.







Оценили 17 человек
30 кармы