

1.9. Кроме того, "мусорная" ДНК могла вообще никак не использоваться. Наличие этой "мусорной" ДНК, вирусов, ретровирусов, транспозонов и проч., происходящие на этом уровне процессы и явления указывают на то, что ДНК и прочие подобные молекулярные структуры существуют не ради организма, а "ради самих себя". ДНК копируются, размножаются, аллели генов конкурируют друг с другом за локусы, транспозоны "перепрыгивают" с места на место в ДНК, ретротранспозоны копируют себя и вставляют копии в другие участки ДНК, и т.д. Организмы существуют лишь для помощи ДНК в этом процессе, служат транспортными средствами и инструментами для копирования и размножения. ДНК (или их кусочки – гены), создающие вокруг себя более приспособленные к окружающей среде организмы, в силу простой статистики, размножаются и распространяются эффективнее. Но ДНК не обязаны размножаться и конкурировать друг с другом только таким путём. Это может происходить и внутри одного организма. Раз в клетке есть аппарат для копирования ДНК, то она и будет копироваться, независимо от того, оказывает это воздействие на фенотип организма или нет. Это объясняет наличие "лишней" ДНК. Организм для ДНК, а не ДНК для организма. [Р. Докинз. Расширенный фенотип. М.: Астрель: CORPUS, 2011] Это опровергает любые подозрения на какой-то разумный замысел, желавший создать человека.
1.10. Живые организмы, включая человека, носят в себе огромное количество "генетического мусора": "мусорная" ДНК, всякие генетические рудименты, ретровирусные вставки и т.д. Живые клетки содержат массу сложных механизмов для ремонта и копирования ДНК, производят и "носят в себе" множество нужных для этого белков. В том числе и для "обслуживания" всего этого "мусора", составляющего большую часть нашего генома. Клетка тратит на это много ресурсов. Это одно из проявлений нерационального устройства организмов (наряду с многими анатомическими нерациональностями, рудиментами и прочими нерациональностями в природе, о которых говорится ниже). Это указывает на отсутствие разумного создателя и на "слепую" эволюцию. Помимо этого, многое из этого "генетического мусора" (рудименты, ретровирусные вставки и проч.), как указывалось выше, указывает на наше родство с другими животными или на происхождение от них.
1.11. Всё это позволяет создать так называемые молекулярные часы. Суть их состоит в том, что у каждого гена есть своя характерная скорость (или частота) мутации. Это позволяет по измерениям гена проследить его состояние в прошлом и измерить промежуток времени, прошедший с момента, когда он имел какое-то другое состояние. Так можно датировать возраст более ранних видов животных и растений. Если, в какой-то момент времени в прошлом геномы двух видов становятся одинаковыми, то это точка разветвления этих двух видов (геном их общего предка). Момент времени этого разветвления, как ясно из вышесказанного, легко определить. Такие методы датировки дают согласованные результаты с другими методами (например, разными радиометрическими методами, методом аминокислотной рацемизации и др.).
2. Морфологические доказательства
2.1. Генеалогическое древо. Аналогичное генеалогическое древо можно построить и на основе изучения фенотипов животных, то есть их анатомического строения (сравнительная анатомия) и физиологии. Этим занимается целая отрасль биологической науки, называемая таксономией, которая посвящена классификации живых существ. Биологи-исследователи, называемые фенетиками (числовыми систематиками, измерителями чистого подобия или измерителями среднего расстояния) измеряют все фенотипические признаки живых существ и строят многомерное пространство, где по осям координат откладываются числовые характеристики этих признаков. Каждый вид изобразится точкой в этом пространстве. Далее вычисляются средние расстояния между этими точками, находятся кластеры, сгущения, иерархия, вычисляется индекс среднего подобия и т.д. В результате строится генеалогическое древо живых существ. И оно совпадает с упомянутым выше древом, построенным на основе молекулярной биологии.
При делении организмов по группам (домены, надцарства, царства, типы, классы, отряды, семейства, роды, виды и пр.) группы более низкого уровня полностью входят (объединяются) в группы более высокого уровня, без каких-либо пересечений и наложений ("принцип полной вложенности"). Это указывает на естественность этой классификации и отражает ветвление эволюционного древа. [Р. Докинз. Слепой часовщик].
Сейчас это генеалогическое древо уже является общепризнанным среди биологов. Его можно увидеть, например, здесь. Это сокращённый вариант, так как здесь всего 3000 видов, а всего их на Земле – десятки миллионов. Например, пункт "Homo Sapiens" включает здесь в себя не только человека, а всех приматов.
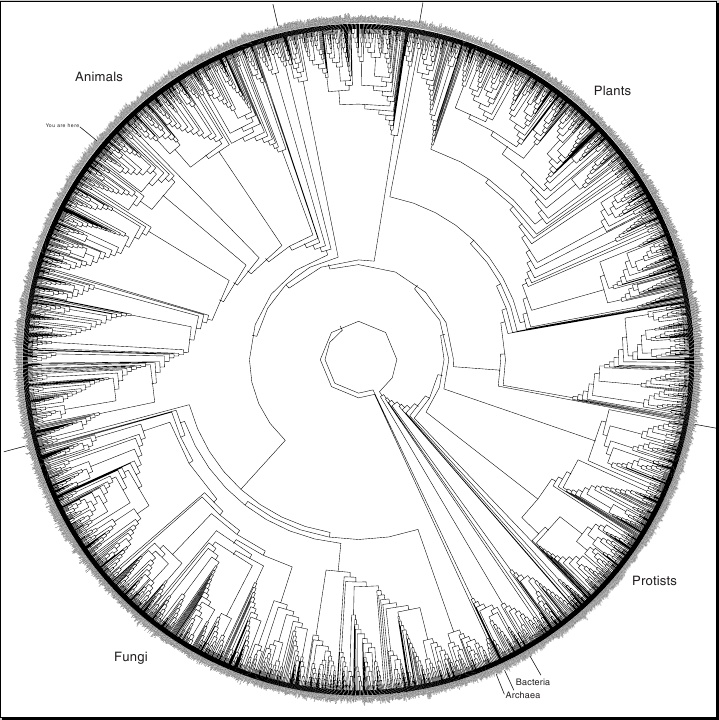
На первый взгляд может показаться, что совпадение молекулярно-генетического и морфологического деревьев – это "тавтология", поскольку фенотип определяется генотипом, а значит, они и должны всегда совпадать. Однако это не так. (1) Помимо генов, кодирующих белки, или генов, регулирующих их работу, в геноме есть множество всякого "мусора", который не влияет на фенотип, но в котором тоже происходят мутации, и со временем накапливаются различия после разделения двух филогенетических ветвей. (2) Каждую конкретную аминокислоту можно закодировать несколькими разными кодонами, и это никак не скажется на фенотипе. Хотя с точки зрения морфологии человек и шимпанзе несравненно ближе, чем человек и мышь, на генном уровне можно было бы сделать человека больше похожим на мышь, чем на шимпанзе. Для этого надо было бы у человека и мыши одинаковые аминокислоты в соответствующих белках (например, в гемоглобине) кодировать одинаковыми кодонами, а у шимпанзе – каким-нибудь другим. И так по всему геному. Также можно было бы накидать огромное количество одинаковых мутаций в "мусорную" часть ДНК мыши и человека, которых нет у шимпанзе. Это никак не сказалось бы на морфологии, физиологии, поведении, умственных способностях и т.д. этих видов. Другими словами, с точки зрения творения генетическое и морфологическое древо совсем не должны были бы совпадать. Поэтому их совпадение является доказательством эволюции, родственной связи видов, объективного существования филогенетического древа.
2.2. Общность строения тел. Чем более схоже организмы, тем ближе их родственная связь. Например, скелеты позвоночных животных (рептилий, птиц, млекопитающих, включая человека, и т.д.) полностью гомологичны (гомеоморфны), то есть состоят из одной и той же комбинации костей. Различаются только сами кости размером и формой. Шотландский зоолог Дарси Томпсон показал, что скелет любого позвоночного можно превратить в скелет любого другого позвоночного путём известного в математике отображения, называемого гомеоморфизмом, и представляющего собой, грубо говоря, растягивание или сжатие тех или иных его частей в разных пропорциях. Аналогичное справедливо не только для скелетов позвоночных, но, например, экзоскелетов ракообразных и т.д.(В какой-то степени это верно и для прочих систем органов - пищеварительной, дыхательной, кровеносной и др.) Палеонтолог и профессор анатомии Нил Шубин пишет: "Лучшая карта человеческого тела – в телах других животных… Это потому, что тела этих существ часто представляют собой упрощённые версии наших собственных тел" [Нил Шубин. Внутренняя рыба: История человеческого тела с древнейших времён до наших дней. М.: Астрель: CORPUS, 2012. С.9-10]. Это явное указание на родственность живых организмов, происхождение от общего предка.По мере эволюции заложенная в генах программа эмбрионального развития удлиняла одни части костей, укорачивала другие части, изменяя у разных потомственных линий размер и форму костей, оставляя неизменным общий план строения скелета (архетип), доставшийся от общего предка. Например, вышедшие на сушу древние кистепёрые рыбы положили начало всем наземным животным на рубеже девонского и карбонового периодов около 360 миллионов лет назад. И скелеты их плавников гомологичны скелету наземной пятипалой конечности (эти рыбы обзавелись также лёгкими, малым кругом кровообращения и 3-камерным сердцем). [Р. Докинз. Самое грандиозное шоу на Земле. Доказательства эволюции. М.: Астрель: CORPUS, 2012.]
Ещё один из таких фактов общности – это сегментированное строение тела. Оно очевидно у многих современных животных, например, у членистоногих, у ланцетника, а также у многих ископаемых. Однако его остатки отчётливо наблюдаются в строении, например, скелета (позвоночника, рёбер...), кровеносной и нервной систем (например, пара нервных волокон, выходящих из каждого позвонка) у млекопитающих, включая человека, указывая на происхождение от сегментированных животных. У эмбрионов млекопитающих она проявляется и в наличии жаберных дуг. Эта сегментированность оставила множество следов в теле млекопитающих, в том числе и такие, которые создают лишние сложности и запутанности, противоречащие "разумному дизайну". Например, см. ниже про возвратный гортанный нерв и про сеть черепно-мозговых нервов. Следы сегментированности есть даже в строении черепа, расположении составляющих его костей (хотя и сильно "затёрлись").
2.3. Неразумный дизайн. Согласно гипотезе о разумном замысле, организмы должны быть устроены не по принципу "лишь бы работало", а аккуратно, короче говоря, разумно, как японская техника с аккуратными спайками, аккуратно уложенными проводочками и т.п. Не должно быть ничего лишнего. На самом деле, организмы как раз устроены по принципу "лишь бы работало", включая бесформенные комки и сплетения нервов, кровеносных сосудов, лимфатических сосудов и узлов и т.д.
Возвратный гортанный нерв отходит от мозга вниз (в составе блуждающего), огибает аорту, а потом возвращается наверх к гортани. У жирафа этот путь достигает 4 метров, хотя от мозга до гортани несколько сантиметров. В этом нет никакой нужды для организма. Более того, из-за этого возникает икота. Ещё пример – расположение семявыводящих протоков, огибающих мочевые каналы, идущие от почек к мочевому пузырю. Это опровергает гипотезу разумного замысла. Зато доказывает теорию эволюции, движимой естественным отбором, который действует "вслепую", и которому действительно не важно, рационально или нет, аккуратно или нет, лишь бы работало. В частности, путь возвратного гортанного нерва становится вполне понятным, если вспомнить, что млекопитающие произошли от рыб. У них нет шеи, и расположение этого нерва (как и других нервов) вполне естественно переплетается с кровеносной системой. В ходе дальнейшей эволюции аорта сдвинулась намного дальше от головы. Но для того, чтобы нерв "разорвался" и "склеился" (условно говоря) с другой стороны аорты, т.е. пошёл в обход, требовалась бы слишком маловероятная мутация. Эволюция пошла по пути постепенного удлинения нерва с сохранением того же расположения относительно аорты. [Р. Докинз. Самое грандиозное шоу на Земле. Доказательства эволюции. М.: Астрель: CORPUS, 2012.]
Эволюция глаза. Многие животные обладают крайне примитивными и несовершенными глазами, которым далеко до глаза млекопитающих. Во-первых, они являются примерами неразумного дизайна. Во-вторых, они наглядно демонстрируют, что глаз не является "неделимой сложностью" и мог эволюционировать от более примитивных форм. Ланцетник воспринимает свет при помощи специального углубления, выстланного светочувствительными клетками. Они объединены в сеть подобно клеткам сетчатки позвоночных, и присоединены к переднему концу нервного тяжа – примерно так же, как наши глаза к мозгу. За формирование этих клеток отвечают те же гены, что формируют и глаза позвоночных животных, включая человека (см. ниже). У морского блюдечка имеется совсем примитивный глаз – просто углубление со слоем светочувствительных клеток. У морского животного Наутилуса глаз чуть-чуть посложнее - он тоже не имеет ни роговицы, ни радужной оболочки, ни хрусталика, а представляет собой просто отверстие, в котором изображение строится по принципу камеры-обскуры. У морского гребешка глаза имеют зачаток хрусталика. Но нет зрительного центра в мозгу. Его примитивные глаза позволяют различать свет и тьму и ощущать движение хищников. Какое разочарование для креационистов, считающих, что глаз не способен эволюционировать от более примитивных форм.
Кстати, светоулавливающие молекулы опсина в светочувствительных клетках наших глаз очень похожи на некоторые молекулярные структуры бактерий. В 2001 г. Детлев Арендт обнаружил, что у многощетинковых кольчатых червей под покровами тела содержатся светочувствительные выросты нервной системы с щетинками, очень похожими на выросты наших палочек и колбочек, и содержащие опсины, близкие к опсинам позвоночных. А как показали генетические эксперименты Вальтера Геринга, за формирование глаз у разных животных отвечает один и тот же ген Pax 6, который можно пересаживать даже от насекомого к млекопитающему, и он будет работать. Случайно ли всё это? [Нил Шубин. Внутренняя рыба. История человеческого тела от древнейших времён до наших дней. М.: Астрель: CORPUS, 2012]
Ген Pax-6 участвует в формировании глаз у человека, мыши, дрозофилы, кальмаров, червей и т.д. Также глаза всех этих животных состоят из одних и тех же типов клеток – фоторецепторных и пигментных (несмотря на то что сами глаза могут иметь разное строение). Все фоторецепторные клетки регистрируют свет с помощью белков-опсинов. Это означает, что все эти группы животных унаследовали ген Pax-6, опсины и эти типы клеток от далёкого общего предка. Впоследствии на основании его (возможно, примитивных) глаз в ходе разных эволюционных путей развилось всё многообразие глаз вышеназванных животных. "С помощью компьютерного моделирования Дан Нильсон и Сюзанна Пелгер из Университета Лунда (Швеция) показали, что отбор малых вариаций за 2000 стадий на протяжении 500,000 лет мог привести к образованию камерного глаза из простого прототипа". [Кэрролл, Шон. Приспособиться и выжить! ДНК как летопись эволюции. М.: АСТ:CORPUS, 2015. Глава 8.]
Инвертированная сетчатка. Ещё пример неразумного дизайна: у позвоночных сетчатка глаза стоит задом наперёд. Это опровергает "разумный замысел" и в то же время доказывает происхождение глаза путём эволюции от более примитивного общего предка хордовых. Сетчатка состоит из нескольких слоёв нейронов, и фоторецепторы (палочки и колбочки) находятся не с той стороны, откуда падает свет (т.е. с внутренней, примыкающей к стекловидному телу), а с противоположной (внешней, примыкающей к эпителию). Сами зрительные нейроны расположены с той стороны от фоторецептора,откуда падает свет. [см.]. И световым лучам, чтобы добраться до рецепторов, приходится продираться через несколько слоёв нейронов (горизонтальных, биполярных, амакриновых, ганглионарых), через сплетение нервов (плексиформные слои) и кровеносных сосудов, из-за чего свет сильно ослабляется, изображение искажается. Затем, нервные сигналы от рецепторов возвращаются назад, в ту же сторону, откуда пришёл свет, к внутренней стороне сетчатки. Там аксоны зрительных нейронов собираются в один пучок посередине сетчатки и уходят за неё, снова пересекая все эти слои зрительных нейронов и капилляров и образуя в этом месте слепое пятно. Ну полная халтура! В результате глазу приходится совершать микроскопические движения, мозгу приходится тратить много ресурсов на очистку изображения от этих помех на основе обработки множества таких нечётких картинок, а также заполнять "дырку" от слепого пятна "заплаткой" на основе изображения, полученного от второго глаза. А мозг и так обходится организму слишком дорого (большой мозг увеличивает смертность при родах и потребляет 20% калорий). Если бы сетчатка стояла бы как надо (как у беспозвоночных), можно было бы или чуть уменьшить мозг, или использовать освободившийся ресурс для чего-то другого.
Зато такое нерациональное устройство сетчатки объясняется эволюцией от относительно примитивного общего предка хордовых, включая позвоночных и ланцетника (относящегося к головохордовым). Ланцетник – довольно примитивное животное, более всего похожее на этого предка хордовых из всех ныне живущих существ.
У самых примитивных многоклеточных расположение рецепторов было устроено довольно логично: рецепторы снаружи внешнего покрова, а питание к ним подводится изнутри. У нынешних беспозвоночных (например, головоногих моллюсков) глаза развивались как впячивания поверхности, поэтому у них сетчатка не инвертирована, фоторецепторы расположены снаружи эктодермы (т.е. направлены внутрь глаза, в ту сторону, откуда падает свет), а аксоны и кровеносные сосуды находятся с внутренней части эктодермы (или с наружной стороны глаза). У хордовых эволюция пошла иначе. Образовавшаяся у них позднее нервная система, как известно, в ходе эмбрионального развития формируется из внешнего зародышевого листка (эктодермы) путём его впячивания и "вворачивания" внутрь самого себя. Получаются две трубки – внешняя (из которой формируются внешние покровы, например, кожа) и внутренняя (из которой формируется нервная система). В результате в нервной трубке рецепторы оказываются внутри, а кровеносные сосуды и нервы подводятся снаружи.
Нервная система ланцетника представляет собой полую тонкостенную нервную трубку с полым утолщением на переднем конце (у позвоночных она развилась в спинной и головной мозг соответственно). Реснички нейронов направлены внутрь трубки. На переднем конце трубки имеются нейроны, играющие роль светочувствительного пятна-глаза. Реснички этих нейронов (которые у позвоночных развились в улавливающие свет палочки и колбочки) также обращены внутрь трубки (т.е. в сторону, противоположную той, откуда падает свет). Ему это не мешает видеть в силу прозрачности тела. Зато такое расположение фоторецепторов более безопасно для них. И ланцетник, и позвоночные унаследовали такое строение от их общего предка.
Как известно, часто (хоть и не всегда и не во всём) развитие в онтогенезе повторяет развитие в филогенезе. Мозг у эмбриона позвоночного животного (включая человека) тоже поначалу формируется как полая трубка с обращёнными внутрь нервными клетками. На переднем конце трубки образуются два выроста, а на их кончиках формируются углубления (это будущие глаза), на внутренней поверхности которого размещаются нейроны сетчатки, по-прежнему обращённые нервными окончаниями внутрь нервной трубки (или наружу по отношению к глазу с задней его стороны – в сторону, противоположную той, откуда падает свет). А аксоны зрительных нейронов и капилляры оказываются снаружи нервной трубки, т.е. на внутренней поверхности глаза. Так позвоночные животные, включая человека, унаследовали инвертированную сетчатку от ланцетникоподобного предка, жившего около 500 миллионов лет назад (в Кембрийском периоде). Его пятно из светочувствительных клеток эволюционировало сначала в пару фотодетекторов в форме чашевидных углублений, позволяющих захватывать больше света (и чуть лучше определять направление на источник). В ходе эволюции постепенно углубление замкнулось в сферический глаз, способный формировать на сетчатке изображение. [Карл Циммер. Эволюция: Триумф идеи. М.: Альпина нон-фикшн, 2013. С. 210-214]
Некоторые креационисты утверждают, будто инвертированность сетчатки задумана для теплоотвода – мол, если бы сетчатка наземных позвоночных была неинвертированной, она перегрелась бы. А вот у осьминогов или кальмаров, живущих под водой, сетчатка неинвертированна, потому что там мало солнечного света, и опасности перегрева нет. Это мнение не выдерживает критики, уже хотя бы потому, что некоторые беспозвоночные проводят много времени на суше или живут возле поверхности пресноводных рек, и отчего-то сетчатка у них не перегревается. В то же время существует множество глубоководных позвоночных (например, рыбы), которые живут в тех же условиях и при той же освещённости, что и подводные беспозвоночные (например, кальмары), но у них сетчатка инвертирована. Зачем? Вообще, позвоночные и беспозвоночные не разделены по разным условиям обитания, а живут вперемешку друг с другом, как на суше, так и под водой. Очевидно, что инвертированность или неинвертированность сетчатки не связана с образом жизни, а связана с происхождением, с эволюционным развитием. Филогенетические линии беспозвоночных разошлись с линией хордовых раньше, и они не унаследовали такое строение сетчатки. Тем более, моллюски и прочие беспозвоночные обзавелись панцирями и раковинами раньше, чем глазами. И поскольку они, в отличие от ланцетника, потеряли прозрачность, у них фоторецепторы и не могли развиваться внутри нервной трубки. [См. "История глаза"]
Эволюция уха. Анатом и палеонтолог Нил Шубин также ясно и доказательно описал в книге "Внутренняя рыба" и эволюцию уха. Начнём с внутреннего уха. Древнейшее известное позвоночное имело всего 1 полукружный канал. У более поздних организмов их 2, а у большинства современных рыб и других позвоночных – 3. Орган боковой линии у рыб по строению и функционированию очень похож на внутреннее ухо млекопитающих и рептилий: и то, и другое представляет собой полость, заполненную желеобразной жидкостью, внутрь которой проходят волоски нервных клеток. Когда эта жидкость смещается, волоски отклоняются, и нервные клетки посылают в мозг сигнал. Но у рыб этот орган более примитивный, им он позволяет улавливать течение воды возле себя, а млекопитающим (включая человека), амфибиям и рептилиям – воспринимать звуковые сигналы и ощущать положение и ускорение своего тела. А эти самые волосковидные нервные клетки есть и у ланцетника, но выполняют иную функцию, то есть они появились задолго от образования ушей. У амфибий и рептилий внутреннее ухо устроено проще, чем у млекопитающих и не завито наподобие улитки. Почему создатель о них не позаботился? Потому что это эволюция, а не разумное творение. За формирование органа боковой линии у рыб и внутреннего уха у амфибий, рептилий и млекопитающих отвечает один и тот же ген – Pax 2. Это говорит о том, что у этих органов общая история. Ген, представляющий собой мозаичную смесь гена Pax 2, отвечающего за формирование внутреннего уха, и гена Pax 6, отвечающего за формирование глаз, имеется у медуз, указывая на нашу связь с ними. Видимо, уши и глаза развились от одних и тех же древнейших структур. Поэтому у человека многие врождённые дефекты сказываются на обоих этих органах.
Кости среднего уха млекопитающих произошли от костей и хрящей рыбьих челюстей. У рыб, амфибий и рептилий челюсти состоят из нескольких костей. При этом среднего уха у рыб нет вообще, а у рептилий оно состоит лишь из одной кости – стремечка (а почему бог не дал им все три?). У акул верхняя челюсть крепится к черепной коробке с помощью хряща – подвеска. Для слуха в воде и в воздухе (на суше) требуются разные органы, поэтому после выхода на сушу у древних амфибий этот хрящ стал менять положение и функцию, превратившись у рептилий в слуховую кость – стремечко. В ходе дальнейшей эволюции рептилий в млекопитающих кости нижней челюсти (кроме одной) сместились во внутреннее ухо и стали молоточком и наковальней. Некоторые другие хрящи нижней челюсти акул у млекопитающих превратились в гортань и некоторые другие структуры горла. Кости, управляющие у акул движением челюстей, у нас в процессе эволюции, движимой естественным отбором, превратились в слуховые кости и кости горла, помогающие нам слышать, глотать и говорить (и они гомологичны им – см. выше).
Всё вышесказанное доказывается:
1) общностью генов, отвечающих за формирование данных структур у рыб, амфибий, рептилий и млекопитающих;
2) формированием этих костей у разных таксонов из одних и тех же жаберных дуг и более поздних структур эмбриона в процессе эмбрионального развития (у эмбриона млекопитающего будущие молоточек и наковальня примыкают к нижней челюсти так же, как и будущие кости челюстного сустава у эмбриона рептилий, и неотличимы от них);
3) гомологичностью слуховых костей среднего уха млекопитающих челюстным костям и хрящам рыб и рептилий;
4) "обслуживанием" одними и теми же нервами этих челюстных костей и хрящей у рыб и рептилий и структур внутреннего уха и горла у млекопитающих (причём это приводит к сильно запутанному и нерациональному переплетению этих нервов, опровергающему разумный замысел, и понятному только с точки зрения "слепой" эволюции – см. ниже);
5) палеонтологическими данными: обнаружено множество скелетов ископаемых переходных форм от рептилий к млекопитающим, у которых хорошо видно постепенное смещение костей челюсти в среднее ухо.
[Нил Шубин. Внутренняя рыба. М.: Астрель: CORPUS, 2012]
Рассмотрим подробнее доказательства 2 и 4. У ланцетника и похожих на него примитивных животных имеется ряд жаберных дуг и щелей, к каждой из которых подходит нерв. Всё довольно просто. У эмбрионов рыб, амфибий, рептилий, птиц и млекопитающих, включая человека, строение похожее, у них имеются жаберные дуги (само по себе – это уже доказательство эволюции) и такая же сравнительно простая их иннервация. Эволюция изменяет анатомию организма, изменяя ход процесса эмбрионального развития. У рыб уже из 1-й и 2-й жаберной дуг эмбриона образуются несколько костей челюстей, а из остальных – развиваются жабры и жаберные щели. У млекопитающих нижняя челюсть состоит из одной кости, а остальные сместились в уши и горло. Из 1-й жаберной дуги, которая иннервируется троичным нервом, образуются верхняя и нижняя челюсти, слуховые кости – молоточек и наковальня – и окружающие их мышцы и сосуды. Поэтому и отростки троичного нерва идут и к челюстям, и к ушам. Из 2-й жаберной дуги образуется третья слуховая кость – стремечко, небольшая кость горла и мимические мышцы. Поэтому отростки лицевого нерва ведут к мышцам лица, к ушам и горлу. Из 3-й и 4-й дуг образуются кости, мышцы, сосуды и нервы горла и гортани. Аналогичное справедливо и для их нервов – языкоглоточного и блуждающего. Всё это и породило очень запутанную и нерациональную сеть. Но несмотря на свою запутанность, система черепно-мозговых нервов человека гомологична системе черепно-мозговых нервов акулы, т.е. каждому нерву человека есть аналог акульего нерва (аналогичное справедливо для нервов всех рыб, амфибий, рептилий, птиц и млекопитающих). И гомологичные нервы ведут к гомологичным структурам головы и горла.
В этой же книге описывается и ещё один пример анатомической нерациональности – это пути черепно-мозговых нервов у человека (и всех млекопитающих): "...Почему два разных нерва, которые иннервируют совершенно разные участки лица и челюсти, посылают ответвления к мышцам уха, лежащим по соседству друг с другом? Ещё больше сбивает с толку то, что троичный и лицевой нервы едва не перекрещиваются, посылая ветви в разные участки нашего лица и челюстей. Почему? Зачем? Функции этих нервов кажутся избыточными, пути – бесцельно запутанными, в их строении не видно ни логики, ни смысла, и совсем уже непонятно, почему те или иные их участки соответствуют тем или иным плитам, блокам и прутьям, из которых состоит наш череп...". [Там же, стр. 115.] Шубин приводит аналогию со старым зданием института, в котором ему приходилось работать. Электропроводка и водоснабжение были проведены столь нерациональным образом и так запутаны, что ни один здравый конструктор не мог спроектировать их таким образом. Это объясняется тем, что здание претерпевало многократные перепланировки и капитальные ремонты. Пути этих нервов также становятся совершенно понятны лишь с точки зрения вышеописанной эволюции, движимой неразумным естественным отбором, которому нет дела до аккуратности.
Во-первых, одинаковые эмбрионы с жаберными дугами, а также все указанные выше доказательства эволюции уха – это доказательства происхождения рыб от примитивных животных, похожих на ланцетника (или от него самого), а млекопитающих (включая человека) – от рыб. Это следы иннервации жаберных дуг ланцетника и рыбьих челюстей (которая намного проще), а также следы сегментированного строения тела (включая и голову), полученного человеком в наследство от более примитивных животных. Кости и хрящи челюстей, ушей и горла человека образовались в конечном итоге из жаберных хрящиков ланцетника. Из относительно простой системы иннервации жаберных дуг ланцетника вследствие изменения процесса эмбрионального развития получилась такая сложная, запутанная и неразумная система иннервации головы и шеи человека. Сегментация "затирается" в ходе эмбрионального развития головы и шеи, но её следы остаются в виде запутанной сети черепно-мозговых нервов и нерационального пути возвратного гортанного нерва. (От лацентника человеку достались и другие проявления сегментированности: например, позвонки и соответствующие каждому из них пары нервов, отходящие от спинного мозга.)
Во-вторых, это явно не разумный дизайн. Запутанные пути этих черепно-мозговых нервов понятны с точки зрения "слепой" эволюции. Это объясняет, почему отростки совершенно разных нервов, выходящих из разных участков мозга, вдруг идут к одним и тем же близким участкам, горла, уха или лица. Вместо того чтобы иннервировать, скажем, все участки уха одним нервом (например, троичным), челюсти и мимические мышцы – вторым (например, лицевым), горло – третьим, и т.д. Разумный создатель так и сделал бы (а не заботился бы о гомологичности нервов млекопитающих, рептилий, амфибий, птиц и рыб). Таким образом, пути нервов головы указывают на "слепую" эволюцию, а никак не на разумного творца.
Ещё пример несовершенств – 2-камерные и 3-камерные сердца. Самое лучшее строение сердца – это 4-камерное сердце, в котором достигается полное разделение венозной и артериальной крови. Разумный творец должен был бы создать такое сердце у всех животных. Но вместо этого мы видим 2-камерные сердца у рыб, 3-камерные у амфибий и 3-камерные с перегородкой у рептилий. Зато млекопитающие и птицы обладают 4-камерными сердцами. Совпадение ли, что у более ранних с эволюционной точки зрения классов наблюдаются более примитивные сердца? Мы здесь наблюдаем не творение разумного замысла, а эволюцию сердца.
Примером "неразумного дизайна" является и совмещение входных путей в пищеварительную и дыхательную системы, что создаёт опасность подавиться. Такое строение наземные животные точно так же унаследовали от рыб, некоторые из которых сотни миллионов лет назад, живя в обеднённых кислородом водоёмах, начали всплывать на поверхность и заглатывать атмосферный воздух. Со временем у них возник вырост на переднем конце пищеварительного тракта, в котором задерживался пузырь воздуха. У одних рыб он постепенно развился в плавательный пузырь, позволяющий им контролировать глубину погружения. У некоторых он начал выполнять по совместительству и дыхательную функцию. Рыбы из надотряда двоякодышащих имеют и жабры, и лёгкие и совмещают жаберное и лёгочное дыхание. Особенно это актуально для рыб, живущих в периодически пересыхающих водоёмах или в водоёмах, обеднённых кислородом. Так, рогозуб, кроме дыхания жабрами, каждые 40–50 минут поднимается к поверхности воды, чтобы заглотнуть атмосферный воздух. Период засухи, когда реки высыхают и мелеют, рогозубы переживают в ямах с сохранившейся водой. Протоптер обитает в пересыхающих водоёмах со стоячей водой и также периодически поднимается к поверхности, чтобы глотнуть воздуха. Молодые особи получают через жабры 90% кислорода, а по мере взросления рыба всё больше полагается на лёгочное дыхание. Взрослые особи 98% кислорода получают через лёгкие. "Арапаима может дышать атмосферным воздухом благодаря ткани пронизанной густой сетью кровеносных сосудов, похожей на лёгочную ткань, которая выстилает глотку и плавательный пузырь, имеющий ячеистое строение и выполняющий функцию дополнительного органа дыхания. Она всплывает к поверхности за воздухом каждые 5–20 минут. Это приспособление развилось вследствие малого содержания кислорода в водах Амазонки".
"Легкие большинства примитивных амфибий (тритонов, амбистом и др.) имеют вид простых мешков, покрытых снаружи капиллярами. Легкие лягушек и жаб имеют внутри складки, увеличивающие дыхательную поверхность. Эволюция дыхательной системы происходила в направлении постепенного расчленения легкого на более мелкие полости, так что строение легких у рептилий, птиц и млекопитающих постепенно усложняется. У ряда рептилий (например, у хамелеона) легкие снабжены придаточными воздушными мешками, которые раздуваются при наполнении воздухом. Животные принимают угрожающий вид – это играет роль защитного приспособления для отпугивания хищников. Легкие птиц также имеют воздушные мешки, распространяющиеся по всему телу... Легкие млекопитающих и человека имеют более сложное и совершенное строение, обеспечивающие достаточное насыщение кислородом всех клеток тела, и тем самым, обеспечивают высокий обмен веществ. Поверхность их органов дыхания во много раз превышает площадь поверхности тела. Совершенный газообмен поддерживает постоянство внутренней среды организма, что дает возможность млекопитающим и человеку обитать в различных климатических условиях". ["Эволюция дыхательной системы" на сайте Evolution.Powernet.ru] Таким образом, даже в современном мире имеется ряд переходных форм дыхательных систем: жабры – сочетание жабр с примитивными дыхательными пузырями у двоякодышащих рыб – дыхательные мешки примитивных амфибий – лёгкие лягушек и жаб с внутренними складками – лёгкие млекопитающих, имеющие губчатое строение, с множеством ветвящихся бронхов и альвеол. Так же происходило развитие лёгких в филогенезе.
2.4. Рудименты также являются свидетельствами "неразумного дизайна" (органы, которые являются "следами" каких-либо органов предковых животных, утративших, сокративших или видоизменивших свою функцию) и атавизмов. Например, копчик – остаток хвоста (и он гомеоморфен ему), пальцы на ногах – рудимент, доставшийся от обезьяноподобных предков, лазавших по деревьям, полулунная складка plica semilunaris во внутреннем уголке глаза – рудимент мигательной перепонки (третьего века) низших позвоночных. Аппендикс – рудимент слепой кишки. Ещё пример – выходные канальца гайморовых и лобных пазух. Почему же бог сделал их сверху, а не снизу? Ему нравится наблюдать за муками людей, болеющих гайморитом и фронтитом? На самом же деле это осталось в наследство от предков, передвигающихся на 4-х лапах. И так далее. Есть ещё множество провизорных органов, проявляющихся у эмбрионов, но исчезающих впоследствии. Иногда люди рождаются с жаберной кистой, с жаберными хрящами, с хвостами или покрытыми шерстью. У китов присутствуют рудименты костей ног, а у змей есть рудименты тазовых костей. Это доказывает, что киты не были созданы такими, какие они есть, а произошли от сухопутных четвероногих. А раз так, то и все существенные полезные адаптации для жизни в воде тоже развились у них в ходе эволюции.
Продолжение. Начало: Часть 1. Часть 2.
Дополнительно:
Атеизм - детям. Гагарин летал - бога не видал.
Атеизм - детям. Эволюция на вашем столе.
Детская энциклопедия в 10 томах
Теория эволюции. Урок 3. Естественный отбор
Теория эволюции. Урок 4. Синтетическая теория эволюции
Зачем нужны гены. Мульт теория
Эволюция - 3. Синтетическая теория эволюции - часть 1
Эволюция от обезьяны к человеку
Жили были первооткрыватели 15 серия. Дарвин и теория эволюции







Оценили 16 человек
27 кармы