

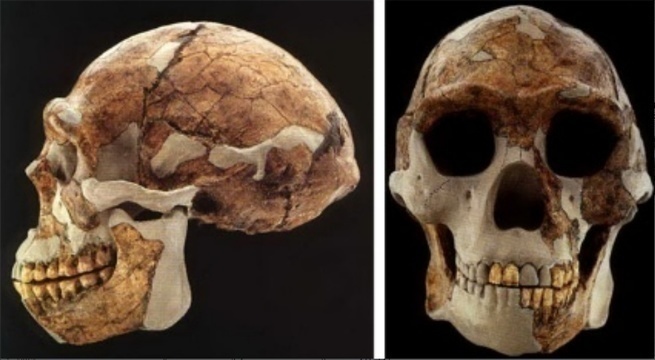
В 1970-х годах большинство учёных пришло к мнению, что птицы произошли непосредственно от определённой ветви специализированных двуногих хищных или всеядных динозавров – Coelurosauria. И действительно, анатомическую связь переходных форм от ящероподобных ранних диапсидных рептилий до современных птиц можно проследить по всей динозавровой линии. Несмотря на то, что осталось несколько противников этой теории, они до сих пор так и не смогли предложить альтернативные модели происхождения птиц и подтвердить их тщательными и многократно повторяемыми исследованиями.
Все исследователи сходятся во мнении, что ближайшими родственниками птиц являются крокодилы; в большинстве случаев эта точка зрения подтверждалась как ископаемыми свидетельствами, так и генетическими доказательствами, указывающими, что крокодилы более тесно связаны с птицами, чем с ящерицами.
Последний общий предок птиц и крокодилов (т.е. предковый архозавр) жил, вероятно, в начале триасового периода на заре мезозойской эры. Вскоре после этого архозавры разделились на две разные ветви – с одной стороны на крокодилов (Pseudosuchia, неправильно называемых «ложными крокодилами»), и с другой стороны на птиц (Ornithosuchia, или «птичьих крокодилов»). Хотя и трудно представить себе две более отличающиеся друг от друга группы животных с точки зрения анатомии, но предков птиц и крокодилов, действительно, объединяли некоторые важные общие характеристики. Например, и те, и другие обладают четырехкамерным сердцем, и, вполне вероятно, что такое же сердце было и у их общего триасового предка. Этот общий предок, возможно, строил гнезда из растительности, а не зарывал яйца в землю, и в некоторой степени демонстрировал заботу о потомстве – ведь даже современные крокодилы защищают свой молодняк от хищников в течение короткого времени после его вылупления. Однако внешне этот предок больше походил на крокодила, чем на птицу. Несмотря на то, что расставленное в стороны положение конечностей и волочащаяся походка современных крокодилов отчасти является результатом их водной специализаций, предок птиц и крокодилов, вероятно, тоже был четвероногим с расставленными в стороны конечностями.
Следующим крупным ответвлением в линии, ведущей к птицам, стал раскол между их предком и птерозаврами – широким спектром мезозойских позвоночных, включающим в себя птеродактилей. Общий предок обеих линий, входивший в группу Ornithodira, вероятно, тоже имел наполовину расставленное в стороны четвероногое положение конечностей и откладывал яйца с кожистой оболочкой, а не с твёрдой и хрупкой скорлупой, как современные птицы. Интересно, что все известные птерозавры имели плотный покров из напоминающих мех волокон – пикнофибр. Вполне возможно, что эти волокна присутствовали и у общего предка птиц и птерозавров, представляя одну из ранних стадий (стадию I, описанную ниже) эволюции перьев. Однако прежде чем мы сможем проверить эту гипотезу, нам предстоит больше узнать о кожных покровах орнитодиров.
У основания линии орнитодиров и непосредственно в подгруппе Dinosauromorpha предки птиц освоили прямохождение. Вероятно, на первых порах это был просто новый способ быстрого передвижения, подобный тому, как при быстрой ходьбе крокодилы принимают более вертикальное положение. У ранних орнитодиров, остававшихся по-прежнему четвероногими, развились очень длинные задние конечности и, вероятно, использовались для бега. У чуть более продвинутых Dinosauriformes все четыре конечности, вероятно, были подведены под тело, а у появившихся вслед за тем настоящих Dinosauria в вертлужной впадине образовалось специализированное отверстие (прободенная вертлужная впадина), постоянно фиксирующее задние конечности в вертикальном положении и делающее невозможным их расхождение в стороны у всех динозавров, за исключением тех продвинутых видов у которых положение задних конечностей было специально адаптировано для плавания и лазания. Первый динозавр, общий предок птиц и орнитисхий (ещё одно неуместное и вводящее в заблуждение название, дословно означающее «птицетазовые»), вероятно, был всеядным двуногим с пятью пальцами кисти (три из которых имели когти) и с пятью пальцами стопы. Вероятно, он жил в начале позднего триаса.
Вскоре после расхождения эволюционных линий предков птиц и орнитисхий у общего предка эузаврисхий (общий предок птиц и завроподных динозавров) стала развиваться птичья дыхательная система, включающая в себя кроме лёгких ещё и так называемые воздушные мешки, проникающие во многие кости скелета и делающие их более «полыми» и воздушными. С появлением группы Theropoda предки птиц ещё более усовершенствовали двуногий и бегающий образ жизни, и вскоре после этого стопа изменилась таким образом, что стала иметь три основных пальца с маленьким четвёртым пальцем, располагавшимся высоко на лодыжке, как прибылой палец, а пятый полностью исчез. Этот маленький прибылой палец сохранится, а затем увеличится и у нескольких групп тероподов видоизменится и будет использоваться при ходьбе, захвате добычи или для усаживания на ветки. Два пальца кисти, лишённые когтей, также будут постепенно редуцироваться, и ко времени появления группы Avetheropoda в кисти останется только три пальца.
В какой-то момент, скорее всего уже после появления аветероподов, представители птичьей линии обзавелись одной из самых важных своих особенностей – оперением. Первые перья были, вероятно, простыми волосоподобными волокнами из бета-кератина, а самые ранние образцы были предположительно (то есть, если они не являются структурами, аналогичными обнаруженным у птерозавров и/или некоторых орнитисхий) определены у примитивного целурозавра Sciurumimus. Самые ранние пуховые перья появляются у группы Tyrannoraptora, причём одновременно у обоих её примитивных представителей (дейнодонтоидов и компсогнатидов), поэтому, скорее всего, это означает, что они фактически появились чуть раньше средней юры, когда жил общий предок двух этих групп. Данные исследований экземпляров орнитомимозавров Dromiceiomimus – группа, которая, вероятно, ответвилась после компсогнатидов, но перед манирапторами в середине юры, указывают на то, что они могли иметь контурные (если не со сплошным опахалом) перья на передней конечности, образующие самые ранние крылья. Чуть более эволюционно продвинутая группа Segnosauria также сохраняет свидетельство наличия относительно длинных перьев на крыльях, однако они являются волокнистыми. Для определения более точной природы этих перьев требуется больше данных, а также лучше сохранившиеся экземпляры представителей этих групп.
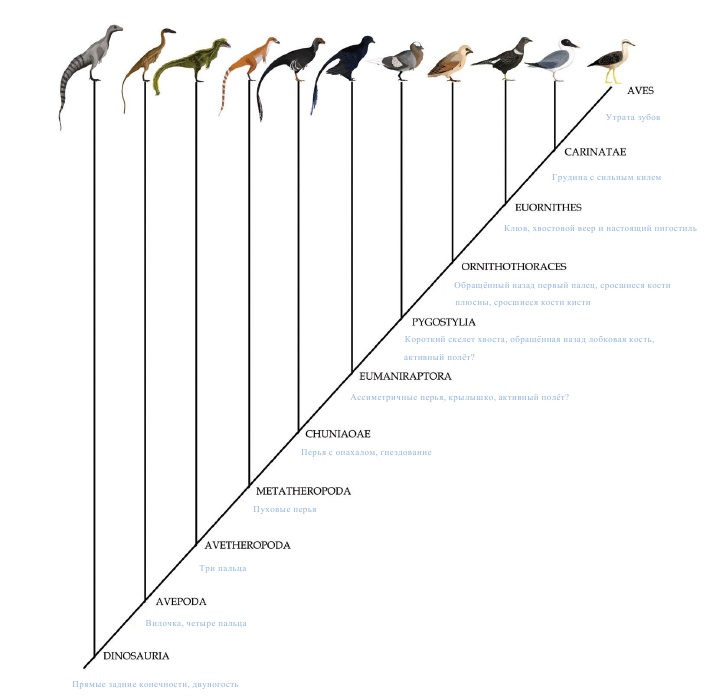
Вскоре после этого у среднеюрского предка птиц и ценагнатиформов появляются полностью современные снабжённые опахалом перья. У этой же группы (Chuniaoae) впервые в летописи окаменелостей встречаются гнёзда для высиживания яиц, вероятно, это связано с превосходной терморегуляционной способностью перьев, имеющих опахало. Затем у группы Eumaniraptora, наряду с перьями, имеющими опахало, появляются асимметричные аэродинамические перья, крепящиеся к малому пальцу кисти, и образуется первое рудиментарное крылышко. Эти адаптации убедительно свидетельствуют о том, что к тому моменту птицы начали предпринимать первые шаги в освоении полёта.
Дальнейшие усовершенствования в анатомии птиц, очевидно, также были направлены в сторону улучшения аэродинамических свойств. К примеру, утрата длинного хвоста случилась до появления группы Pygostylia, после чего развитие адаптаций к полету приобретает уже взрывной характер. У представителей Ornithothoraces изменилась анатомия плеча, что позволило крыльям вытягиваться вертикально и способствовало возникновению полного взмаха при полёте, а большой и малый пальцы кисти срослись воедино, так же, как и кости плюсны. Первый палец из прибылого пальца эуманирапторов, использовавшегося для захвата добычи и лазания, у орнитоторацинов превратился в обращённый назад большой палец стопы, что позволило птицам садиться на мелкие ветки.
Возникшие у Euornithes первые птичьи клювы ещё сосуществовали с зубастыми челюстями, доставшимися этим птицам от предков, а перья хвоста развились в раскрываемую веерообразную структуру, крепящуюся к настоящему пигостилю.
Дальнейшие усовершенствования анатомии крыла позволили взлетать в воздух, исключив необходимость карабкаться на деревья, а когти крыльев значительно редуцировались до размеров когтей Carinatae (хотя маленькие когти до сих пор встречаются у многих современных птиц). Несмотря на то, что киль на нижней стороне грудины появился впервые у представителей Ornithothoraces, у килегрудых он значительно увеличился, что свидетельствует о появлении у них способности к современному полёту, задействующего в основном сильные грудные мышцы. И, наконец, у общего предка всех современных птиц были утрачены зубы, что привело к возникновению характерного беззубого клюва представителей Aves.
Дополнительно:
«Полистратные окаменелости» или «проткнутые слои» с точки зрения геологии. Продолжение.
Мостовая гигантов или столбчатая отдельность.






















Оценили 17 человек
30 кармы